|
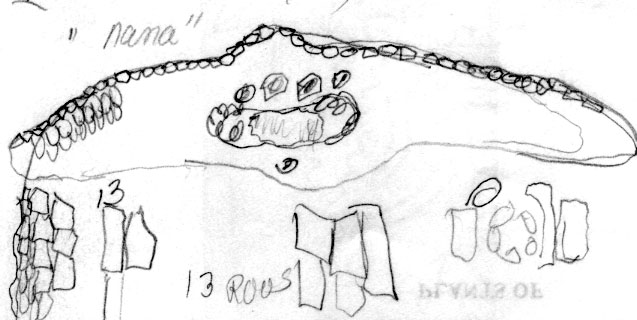
Honshu: Mt. Daisen, Makino 43792 (topotype, S). Illustration indicates abaxial leaf surface lacks papillae across 8 marginal cells across followed by 4 rows of papillose cells, 10–11 stomata rows per band and a midrib 12 cells wide appearing mamillose. |
Honshu: Mt. Daisen, Wilson s.n. (A, topotype). Illustration attached to specimen indicates abaxial leaf surface lacks papillae across 8–9 marginal cells followed by 4 rows of papillose cells, 10–11 stomata rows per band and a midrib 11 cells wide |
Russia—Sakhalin Is.: Schmidt s.n. (GH), with T. umbraculifera var. microcarpa. Illustration indicates abaxial leaf surface lacks papillae across 11 cells, stomata are in 11 rows, and midrib is 16 cells wide, the outer 2 rows with papillae. |
|
Honshu, Hakkada, bush 1–1.5 m, windswept slopes, 1000–2000 m, rare, 4 Jul 1914, Wilson 7133 (A). Illustration attached to specimen shows abaxial leaf lacking papillae across 8 marginal cells, 11 stomata rows per band, and midrib 11 cells wide. |
Honshu: Niigata, Yuzawa-machi, Minamiuonuma-gun, 1650 m, evergreen shrub 0.8 m high, fr red, Taoda 3887 (A). |
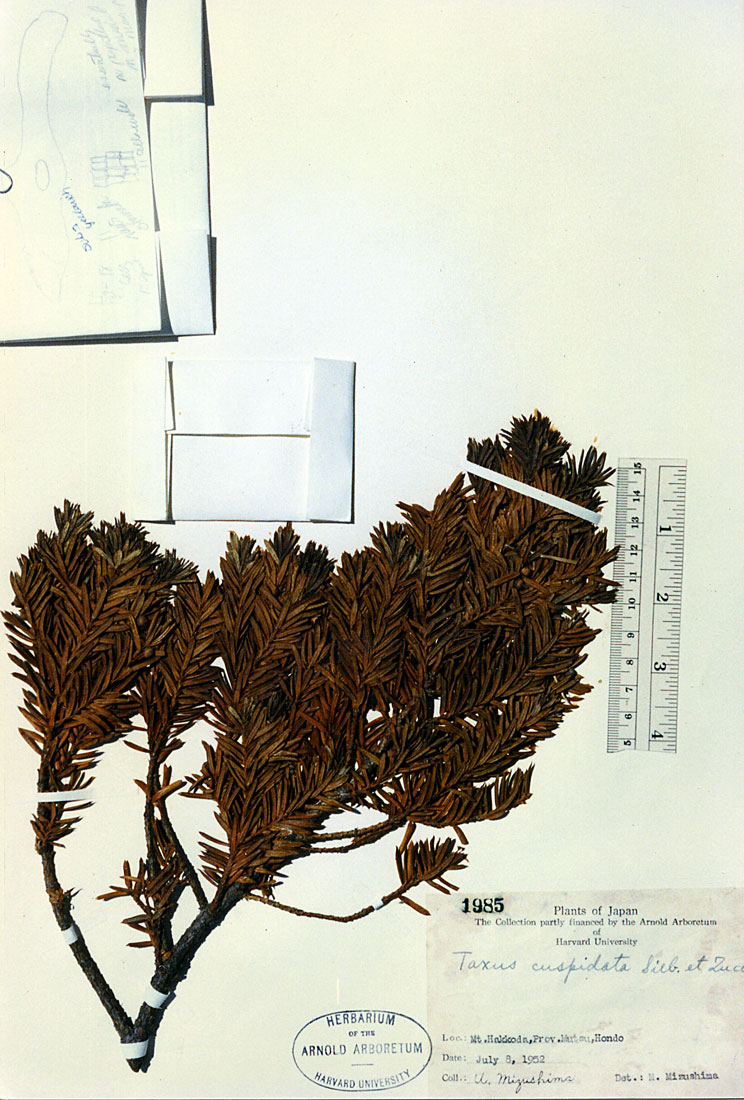
Honshu: Mutsu Prov., Mt. Hakkoda, Mizushima 1985 (A). Illustration attached to specimen indicates abaxial leaf surface has 18 marginal cells (no papillae), 11 stomata tows, and midrib is 11 cells wide (no papillae). |
|
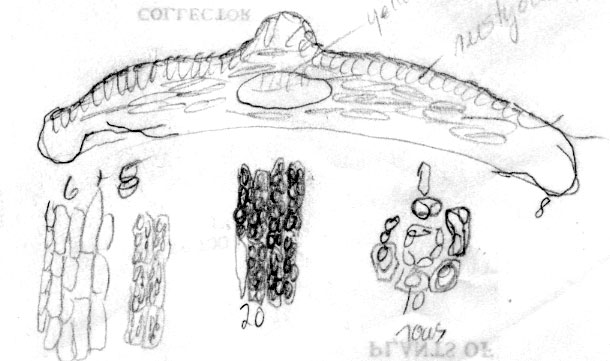
Korea: in forest, Aug 1907, Faurie 1512 (A). Illustration from specimen at BM, shows abaxial leaf margin to lack papillae across 9 marginal cells, stomata 9-10 rows, and a papillose midrib. |
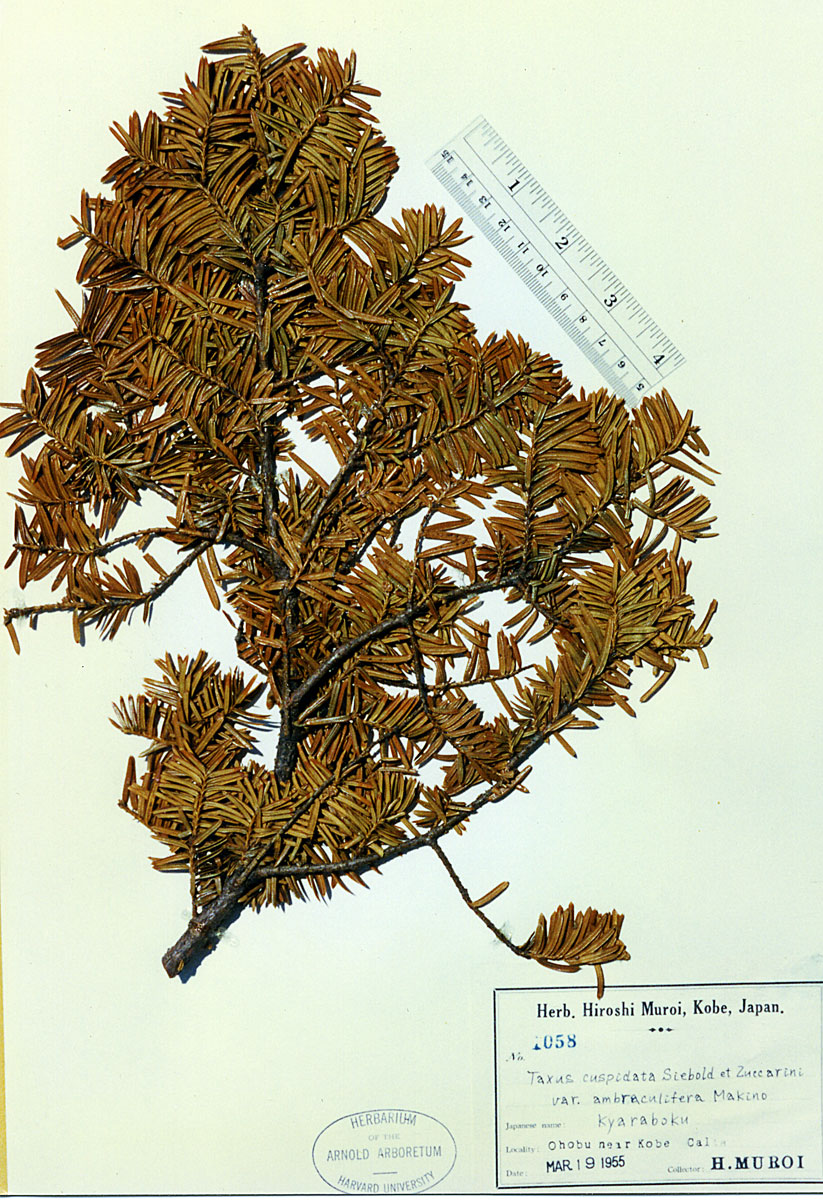
Honshu: Ohobu near Kobe Calta, 19 Mar 1955, Muroi 1058 (A). |
Cultivars: Secrest Arboretum, Wooster, OH. |
|
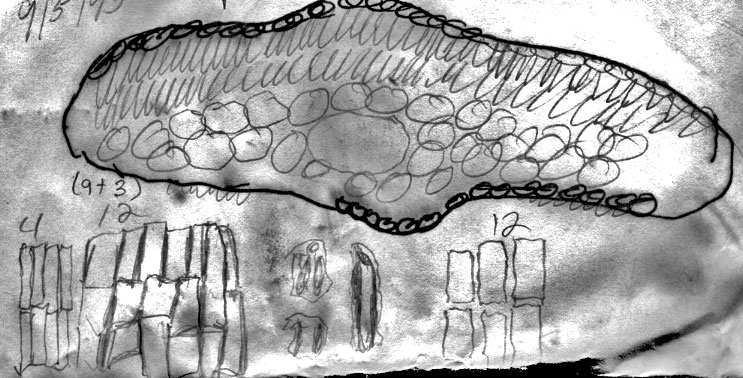
21b. Taxus caespitosa Nakai var. angustifolia Spjut (Figs. 163–164, 271), J. Bot. Res. Inst. Texas 1(1): 268. 2007. Type: Japan. Prov. Kozuke, Oze-ga-hara, “shrubby habit, “leaves dark green,” “yellowish green beneath,” 26 Aug 1950, Mizushima 401 (Figs. 163–164 (holotype A, leaf with a double row of palisade parenchyma cells, lacking papillae across 14 marginal cells, with 9–10 stomata rows/band, seed).
Apparently prostrate bearing numerous erect
reddish orange branchlets, persistent bud-scales few, cuspidate, ~ 1 mm.
long; leaves mostly distichous, spreading from erect branchlets, erect
on horizontal branchlets, reportedly dark green abov Ground cover yew. Korea, Japan.
21c. Taxus caespitosa Nakai var. latifolia (Pilger) Spjut, J. Bot. Res. Inst. Texas 1(1): 269. 2007 (Figs. 159–162, 273–275). Taxus cuspidata var. latifolia Nakai, J. Kor. For. Soc. 158: 39 (1938). Taxus baccata L. [ssp. cuspidata (Siebold & Zuccarini) Pilger] var. latifolia Pilger, Pflanzenreich 4(5): 112 (1903). Taxus cuspidata var. latifolia Nakai, J. Kor. For. Soc. 158: 39 (1938). Syntypes from E Russia, Korea and Japan. Lectotype (designated by Spjut 2007)—Japan: Faurie 6345 at P! (with two labels, one dated 30 Dec. 1890 and another dated 1888, with male cones); isolectotype at K! (fragment). Taxus media Rehder f. hatfieldii Rehder, J. Arnold. Arb.: 198. 1923. Taxus media var. hatfieldii Wilson, Horticulture, n ser. 3: 30, fig. (1925). Type: from horticulture (holotype A!). Broad-leaved yew. Distribution. E Russia, Korea, Japan. Shrubs or trees, variable in habit and branching; main stems prostrate to many spreading upwards from a common base. Branchlets often recurved. Leaves radial on erect branchlets, secund on lateral branchlets, often greenish orange in the herbarium; abaxial marginal cells of (6-) 10–18 smooth cells across, usually with several more quadrate rows of cells near margins, the cells enlarged or rectangular in several or more rows towards stomata bands, 1–3× l/w, 3–5× l/w on midrib, with papillae evident on up to 6 rows of cells, papillose or epapillose on midrib; papillae positioned submarginally in 2 rows on each cell; stomata in 7–13 (-16) rows/band. Seed conical, 3 mm long, 3–4 mm diam., pale yellowish to purplish. Representative Specimens—Russia Federation—Sakhalin Is.: 46º37'N, 142º53'E, Prigarodne, mixed conifer (Abies, Picea)/broadleaved (Betula, Sorbus) woodland, 150 m, to 1.8 m high, Flanakan &Kirkham 203 (K). Korea: in forest, 800 m, 19 Jul 1910, Taquet 4455 (A); without locality data, Faurie 117 (P), 3406 (P), 5975 (P). Japan—Hokkaido: Shiribeshi Prov., Shiribeshi-san, branches wide spreading, 1–2 m, common, 1300-2000 m, 27 Jul 1914, Wilson 7265 (A). Honshu: Mutsu Prov., Mt. Hakkoda, erect tree, 8 Jul 1952, Mizushima 1989 (A). without locality data, Faurie 5114 (P: 2 sheets). Locality unknown: Folley s.n., identified T. cuspidata ‘nana’ (K). Variants—Russia—Sakhalin Is.: Dvorakovskia & Bokina s.n. (A). “Mandshuria” SE: ex herb. hort. bot. Petro. yr 1860, Maximowicz (P [Bunge], S:C-2114). Japan—Honshu: Kanagawa Pref., Mt. Imaizumi, Tomitar ex. Makino 43780 (S: C-2120); Honshu, Akita Pref. Makino 43769 (S); Nikko, Shimotsake, Apr 1887 (P); Nikko, May 1888 Shimotsake 446 (US). Hokkaido: Kitami Prov., Aug. 1897, Sapporo Agric. College (PH)
|
||
|
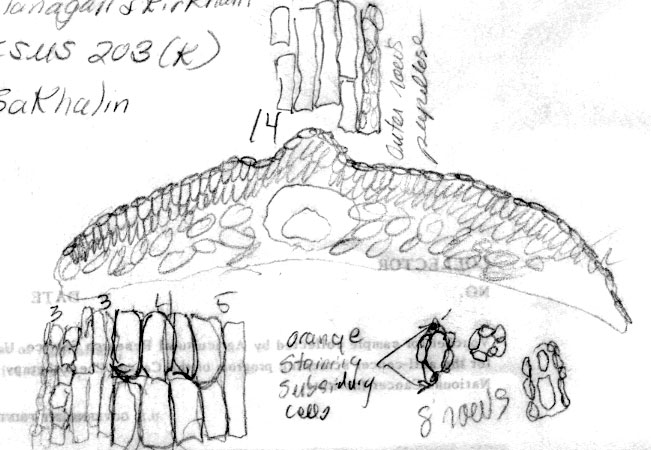
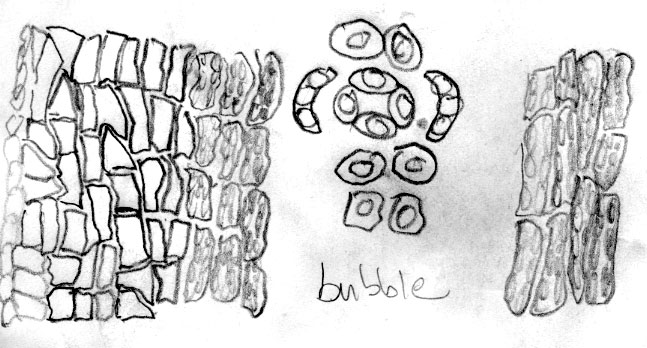
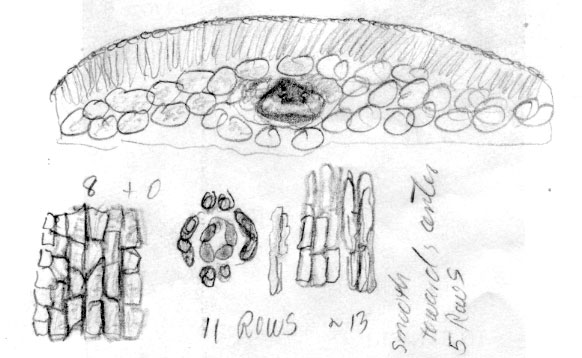
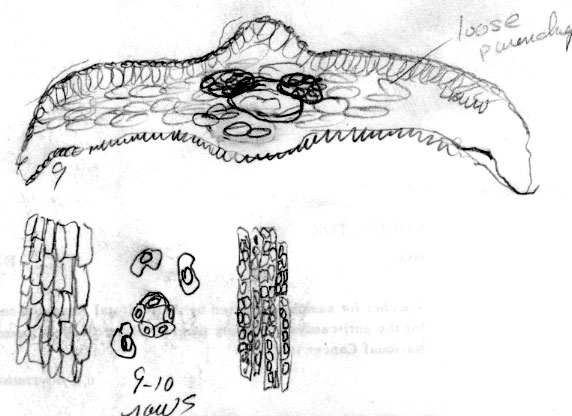
Russia Federation—Sakhalin Is.: 46º37'N, 142º53'E, Prigarodne, mixed conifer (Abies, Picea)/broadleaved (Betula, Sorbus) woodland, 150 m, to 1.8 m high, Flanakan &Kirkham 203 (K). Illustration shows leaf x-section and cell shapes on abaxial surface. The abaxial leaf margin is indicated to lack papillae across 15 cells. This is further shown to be divided into 3 rows of small irregularly quadrate cells nearest the margin, followed by 3 rows of somewhat rectangular cells, 4 rows of inflated cells and 5 rows of more regular rectangular cells nearest the stomata band. The stomata band is indicated to have only 8 rows of stomata. The subsidiary cells were noted to be orange, from which it may be deduced that the specimen was kept in cold conditions before being pressed. Papillae are shown to be low and marginal on accessory cells, and alternate on outer 14 midrib cells. The palisade is a double layer of cells, a feature that is common to the Cuspidata Alliance. |
Korea: Faurie s.n. yr 1910 (A). Stomata rows 13.
Japan: Faurie 5114, yr 1889 (P). Illustration shows leaf abaxial margin to lack papillae across 6 cells, three rows of which are inflated, followed by 4 rows of papillose cells, and 12 rows of stomata. Midrib is noted to be 15 cells wide with obscure medial papillae.
|
Korea: Faurie 3406 (P). Illustration shows abaxial leaf surface to lack papillae across 10 marginal cells of which 3 rows nearest the margin appear relatively small and irregularly quadrate, followed by 7 rows of inflated cells, and 6 rows of papillose cells. The stomata are noted to be in 13 rows. The abaxial midrib is noted to be 14 cells across with low submarginal papillae. The leaf x-section indicates a marginal count of 11 cells across without papillae; thus, the abaxial leaf marginal zone without papillae might be indicated as 10-11 cells wide.
Japan: Nikko, Schimotsake, yr 1880 (P). Annotated T. cuspidata in Oct 1997, but now considered T. caespitosa var. latifolia by the narrow margin of 8 cells, and short epapillose cells on abaxial midrib. |
|
Japan? Faurie 5975 (P). Illustration shows abaxial leaf margin to lack papillae across 10 cells in which 1-2 rows nearest margin are relatively small; this is followed by 7 rows of papillose cells and 7 rows of stomata. Midrib is shown to lack papillae.
|
Cultivated. from Hungary, Folley s.n., identified as Taxus cuspidata nana (K). Illustration indicates abaxial leaf epidermis lacks papillae across 16-17 cells, 1-2 rows nearest margin are relatively small, followed by 2 rows of slightly larger irregularly quadrate cells followed by 13 rows of irregular long hexangular to somewhat rectangular shaped cells, stomata rows are indicated to be 13; midrib cells is shown to lack papillae. The palisade layer is indicated to be double. |
Japan? Faurie 5714 (P). Illustration shows abaxial leaf surface to lack papillae across 14 marginal cells in which two rows nearest margin are short and irregularly quadrate, followed by 2 rows of slightly larger cells, followed by 10 rows of much longer cells; stomata band has 13 stomata rows' midrib has medial papillae. |
|
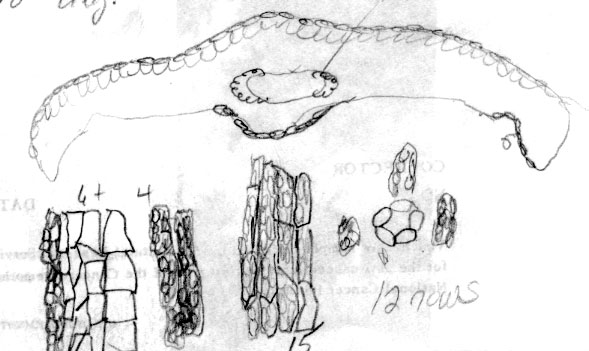
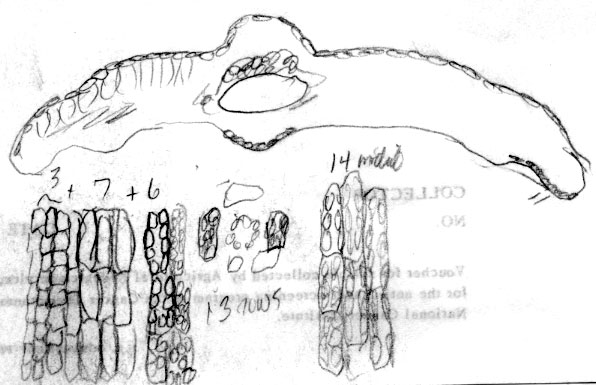
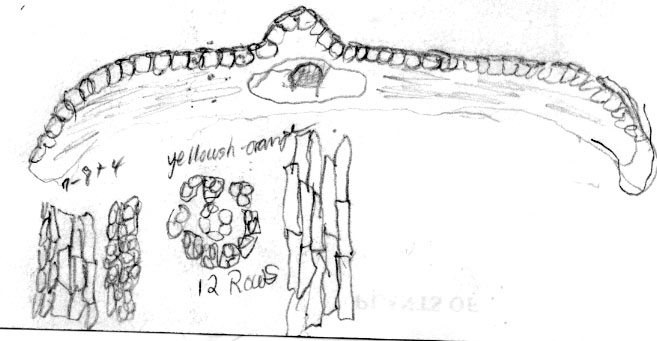
Cultivar: Halorian yew, Secrest Arboretum OH, A30-116. Leaf 2.3 mm wide. Abaxial leaf margin lacks papillae across 16 marginal cells in different size groups (4 + 9 + 3); stomata 12 rows, midrib 12 cells wide, smooth. |
Cultivar: Colean yew, Secrest Arboretum OH, A30-131. Abaxial leaf margin lacks papillae across 8 marginal cells; stomata dense in 11 irregular rows, midrib 15 cells wide, papillose. |
|
|
Erect with Spreading or Ascending Branches |
||
|
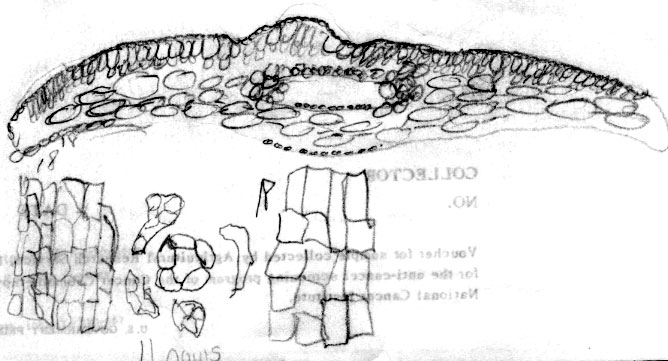
Japan: Faurie 6345, yr 1888. Lectotype (P). Illustration notes abaxial leaf surface lacks papillae across 18 marginal cells, 3 nearest margin are much smaller; there are 11 rows of stomata and the midrib, which is 18 cells wide, lacks papillae. Leaf x-section shows elliptical epidermal cells and double palisade layer, the second rows of cells shorter than the first. Photo on top right shows close-up of male cones; lower right shows stomata band, 250x.
|
Japan, mountains, Faurie 117, yr 1888 (P). Illustration shows abaxial leaf surface to lack papillae across 8 marginal cells, followed by 4 rows of longer obscurely papillose cells, 8 rows of stomata and a papillose midrib 18 cells wide. The parenchyma cells in the leaf mesophyll differ from what is commonly seen. |
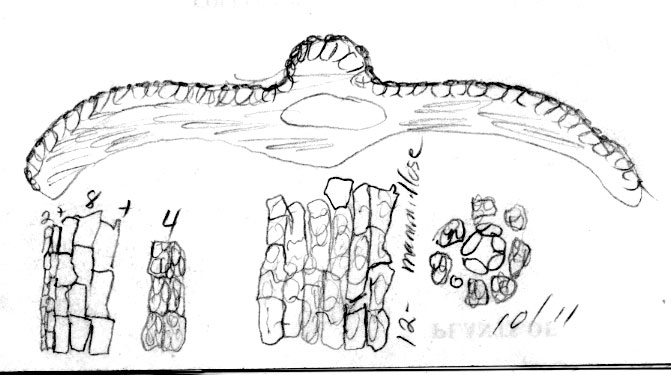
Honshu: Mutsu Prov., Mt. Hakkoda, erect tree, Mizushima 1989 (A). Illustration attached to specimen indicates abaxial leaf surface to lack papillae across 8 marginal cells and also across the entire midrib, which is 16 cells wide. Stomata are noted to occur in 9–10 rows. |
|
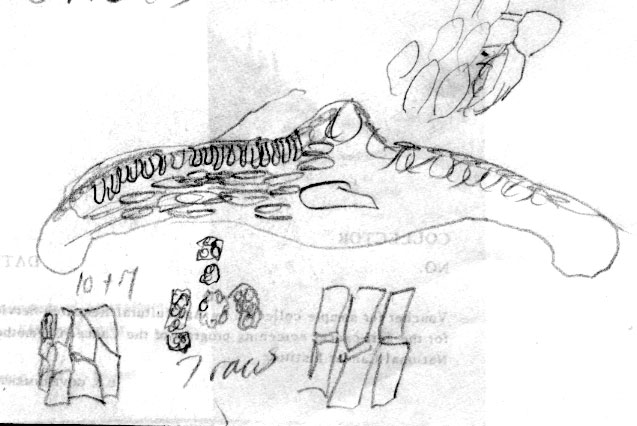
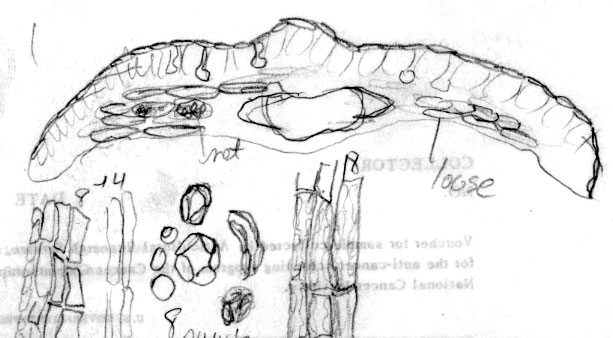
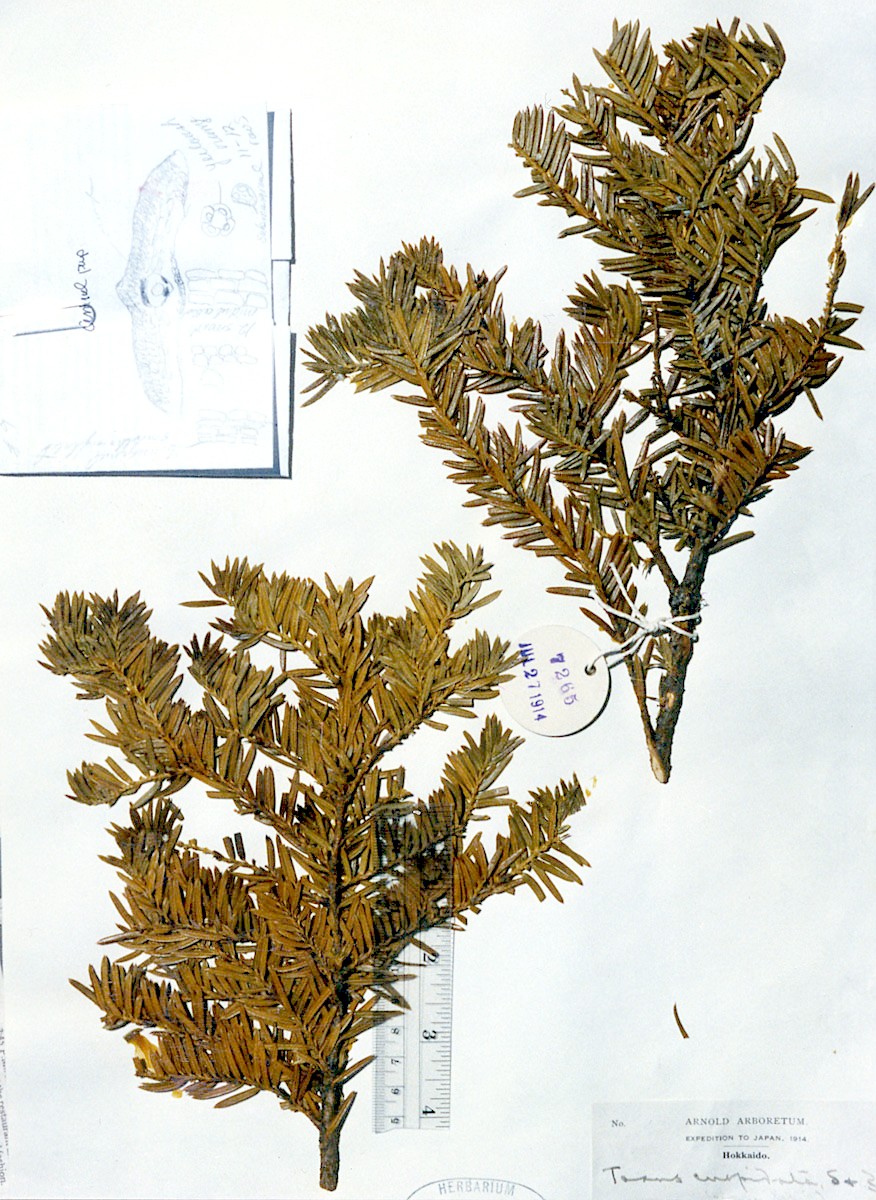
Japan—Hokkaido: Shiribeshi Prov., Shiribeshi-san, branches wide spreading, 1–2 m, common, 1300-2000 m, Wilson 7265 (A). Illustration attached to specimen indicates abaxial leaf surface lacks papillae across 16 marginal cells with two rows of shorter cells nearest margin, and also across the entire midrib, which is 12 cells wide. Stomata are noted to be in 11–12 rows, and surrounded by yellowish orange subsidiary cells. Leaf x-section shows a double palisade layer. |
Honshu: Nikko, May 1888 Shimotsake 446 (US). Leaf sections from this specimen show a strong resemblance to T. mairei var. speciosa; however, the leaf has a thickened lip-like margin as evident in x-section. The abaxial leaf surface lacks papillae across 16 marginal cells in which 5 rows of cells nearest the margin were narrower and longer; stomata were noted to in 15 rows and the abaxial midrib has enlarged epidermal cells lacking papillae entirely, 15 cells wide. |
Hokkaido: Rishiri, Sapporo Agricultural College, yr 1888 (PH). Abaxial leaf margin was noted to be 11 cells across without papillae, stomata in 11 rows and midrib of 14 cells wide was papillose on outer 4 rows.
Korea: Mt. Halla, close-up of midrib cells, 250x |
|
Intermediates to Taxus baccata and T. umbraculifera |
||
|
Russian Federation—Sakhalin Is.: Dvorakovskia & Bokina s.n. (A). Illustration attached to specimen indicates abaxial leaf surface lacks papillae across 8 marginal cells, followed by 14 rows of papillose cells, 11 rows of stomata, and the midrib 11 cells wide. Leaf in x-section shows a double palisade layer. This specimen was annotated T. fastigiata based on color and leaf arrangement, although these features and also flexuous branchlets compare closely with T. baccata var. glauca. However the double palisade layer, 8 marginal cells, and seed tapering from near base are features that favor classification in the Cuspidata Alliance. |
Honshu: Akita Pref. Makino 43769 (S). Illustration indicates abaxial leaf margin lacks papillae across 7–8 marginal cells; stomata bands are yellowish orange with 12 rows of stomata, and midrib lacks papillae. This specimen has branching similar to T. baccata var. jacksonii whereas the color on leaves and branchlets is close to var. glauca. Also, bud-scales are rather inconspicuous as in T. baccata. However, the relatively wide leaf margin and stomata count support placement in the Cuspidata Alliance. |
Japan—Honshu: Kanagawa Pref., Mt. Imaizumi, Tomitar ex. Makino 43780 (S: C-2120). The long branches suggest T. cuspidata, whereas whorled branching and thick branches suggest T. caespitosa var. caespitosa. Illustration of leaf sections indicates abaxial leaf surface has a margin of only 8 smooth cells (no papillose cells); this is followed by a stomata band with 11 rows of stomata, and partially papillose midrib 13 cells wide, lacking papillae mostly on the center 5 rows. |
|
SE Manchuria: Maximowicz, yr 1860. Illustration shows abaxial leaf margin to lack papillae across 6 marginal cells followed by 8 papillose cells, a midrib of 20 cells wide, densely papillose, and stomata in 10 rows. The thick branchlets and phyllotaxy compare favorably with the preceding specimens, but as noted these show similarities to T. baccata. Here the leaf section characteristics also compare more closely to T. baccata. The geographical location is more northern and western. Additional data are needed to support any determination that this and the other specimens of this intermediate group should be classified under T. baccata. Other specimens from Maximowicz reportedly from this region include T. umbraculifera var. umbraculifera and T. umbraculifera var. microcarpa.
|
Cultivar: Secrest Arboretum, Wooster, OH, appearing to be a hybrid, T. caespitosa var. latifolia x T. umbraculifera |
|


















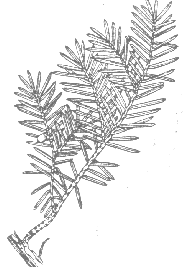
 This
variety is recognized by the relatively thin, narrow leaves as in T.
biternata, but apparently has the habit of T. caespitosa var.
caespitosa as reproduced here from the Illustrated Encyclopedia
of Fauna & Flora of Korea; Vol. 5 (1965).
This
variety is recognized by the relatively thin, narrow leaves as in T.
biternata, but apparently has the habit of T. caespitosa var.
caespitosa as reproduced here from the Illustrated Encyclopedia
of Fauna & Flora of Korea; Vol. 5 (1965).