©The
World Botanical Associates Web Page
Prepared by Richard W. Spjut
April 2003; photos added May 2006; updated August 2007
|
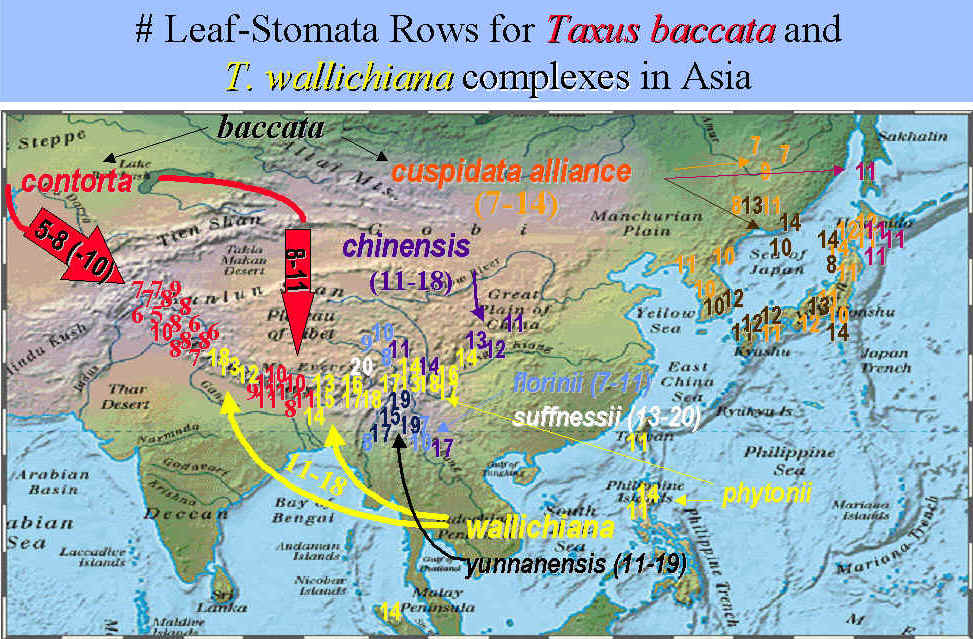
Yunnan: 3000-3200 m, Schneider 2918 (A). See key for illustration of leaf sections. Photos attached to specimen scanned separately and shown below. This photo shows a section of the abaxial surface near mid region of leaf as seen under a microscope, from margin (left), narrow stomata band with 9 rows of stomata (center), and midrib cells with alternate (marginal) papillae on right. Illustration indicates epidermal cells are 40 µm tall and wide. |
Yunnan: Dokerla, 3100 m, Yu 7848 (BM). Temporary annotation label indicated for T. florinii. See key for illustration of leaf sections.
|
  Yunnan: Lung-pan la-Champu, 2800 m, Wang 67414 (A). Illustration indicates leaf along abaxial margin was found with 23 + 5 marginal cells and 10-12 stomata rows per band
|
|
Yunnan: Litiping Range, J.F. Rock 11573 (A). Photos attached to specimen show abaxial leaf sections magnified ~400x. Note marginal papillae. Illustration indicates 10-11 stomata rows per band.
|
Yunnan: S Chungtien, Kung-shiang-shu, Snow Mt, 2700 m, Feng 3235 (A). Photos attached to specimen are of various leaf sections viewed under a microscope at ~250x. Upper left is of midrib cells with papillae aligned along the center and also conspicuous along the margins, upper right is x-section showing large epidermal cells in red; center left is of the vascular bundle; lower right shows part of stomata band, marginal cells with predominantly marginal papillae. |
  Yunnan: Sikang, Me-kong, Tsa-wa-rung, 2500 m, C. W. Wang 65475 (A). Illustration attached to specimen indicates the abaxial leaf surface has 15 marginal cells, 8-9 stomata rows per band, and a midrib of 24 narrow cells.
|
|
Sichuan: 3500 m, Schneider 1429 (K). Illustration indicates abaxial leaf surface has 3 rows of shorter cells followed by 5 rows of longer cells and 15 rows of papillose cells, and then 7 rows of stomata and 22 midrib cells.
|
 
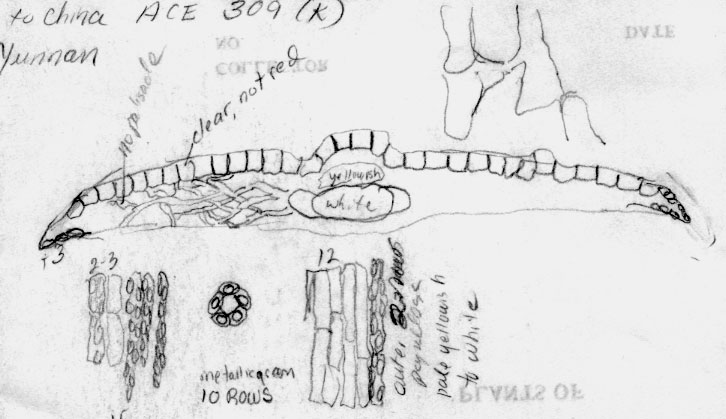
Yunnan: Zhongdian, Haba Shan, 27º22'28 N, 100º05'50 E., 3347 m, Alpine Garden Soc. Exped. 309 (K). Illustration indicates abaxial leaf margin has 2-3 rows of shorter cells followed by 15 rows of papillose cells, stomata band with 10 rows of stomata and a partially papillose midrib, and leaf in x-section did not have reddish epidermal cells as usually seen in T. wallichiana and its allies, but the cells were very large as generally seen for T. florinii. |
|
|
3.
Taxus suffnessii Spjut, J. Bot. Res. Inst.
Texas 1(1): 222. 2007.
Type: MYANMAR. North Triangle (Wring
Tree
or shrub; branchlets unequally divided, gray to purplish on older
growth; bud scales
3–4 seriate, persistent to the 3 rd yr or longer, paleaceous,
indurate, grayish, deltoid, closely to loosely adnate, lower scales
concave with a distinct midnerve, uppermost scales slightly cuspidate
and aristate, ca. 3 mm long and 2 mm wide. Leaves lacking on older
twigs, nearly two-ranked, overlapping slightly, oblong to slightly
elliptical or widest above the mid region, straight, mostly ca. 1.5 cm
long, 3.0 mm wide, 250–350 µm thick, bronze to blackish green and
concave above to a rounded or acutely keeled midrib, yellowish-green and
convex below to a rounded keeled midrib, abruptly revolute near margins.
Upper (adaxial) epidermal cells quadrangular in T-sect., or taller than
wide, to 60 µm tall and 50 µm wide; lower (abaxial) non-stomata
epidermal cells not as large, 12–25 µm high and wide,
quadrangular, papillose to near leaf margins; papillae prominent,
globose, in 3–4 rows, notably medial on marginal cells and marginal on
midrib cells or equally developed on accessory cells in a stellar
arrangement; stomata 12–20 rows/band. Male cones not seen.
Female cones maturing on current season growth, scales overlapping in
5–6 ranks; seed appearing succulent or deformed, conical, dark colored
with notable yellowish neck, ca. 3 mm long, 1.5 mm wide. Suffness yew. Rare, E Himalayas: Endemic to Myanmar. Taxus
suffnessii has perhaps more ancestral features than other species in
the genus.
These include the relatively large bud-scales with a distinct midnerve,
the numerous stomata on leaves—up to 20 rows—in bands that are
scarcely differentiated from adjacent epidermal cells, and the papillose
cells that extend across the midrib and to the margin, or nearly so (0–2
cells from margins). Additionally, its phyllotaxy is most similar to
that of Pseudotaxus (Nothotaxus) chienii, Taxus chinensis and T. brevifolia (Pacific
NW America). Its
antiquity is further evident in comparison to the Pacific yew (T.
brevifolia) by
the similarity in branching, paleaceous bud-scales, and tall angular
leaf epidermal cells. Ironically, Kingdon Ward noted T. suffnessii to
occur “in thickets,” while in this paper T. brevifolia var.
reptaneta is described as a thicket forming yew. The key
difference between these species is that T. suffnessii has more stomata
on its leaves. Its accessory cells also have papillae in
concentric (stellar) patterns rather than aligned in rows. The
bronze-colored leaves—reported by the collector, which are still
retained in dried specimens—are also distinctive in the type. Taxus suffnessii, however, will most likely be difficult to distinguish from T. wallichiana; it differs by the smaller, more polished, nearly nerveless bud-scales, and sometimes by the older reddish orange branchlets as in the typical variety in NE India, in contrast to a purplish color in T. suffnessii. Intermediates may be recognized in specimens collected by Kingdon Ward 21901 from W Central Myanmar and Beer 25316 from Nepal (above Sedua, 9400 ft.). These have slightly larger bud-scales with a midnerve evident above the mid region of the scale. Taxus
suffnessii is named in honor of the late Matthew Suffness in
recognition of his dedication to screening natural products in the
search of new drugs to treat cancer, and the strong encouragement I received from him
in this endeavor. He became Chief of the Natural Products Branch
in the National Cancer Institute after Jonathan Hartwell retired in
1976, following a brief leadership by John Douros. In 1986, Dr.
Suffness became more involved in extramural contracts, often serving as
consultant to various drug discovery groups. He focused on identifying
novel leads that showed promise for development as new anticancer drugs,
one of which was taxol. It is ironic that a major compilation on
taxol research to which he served as sole editor (Suffness 1995) would
not appear until just after his death from cancer in the spring of 1995. Representative specimens: Myanmar: Upper Burma, Hkyet, 27º45' N, 97º50' E, 9000–10,000 ft., Kingdon Ward 13003 (BM); Myintkyina, Sumprabum, 8600 ft., Hla & Koko (K); W Central Esakan, 6400 ft, in thick forest, Kingdon Ward 21901 (A).
|