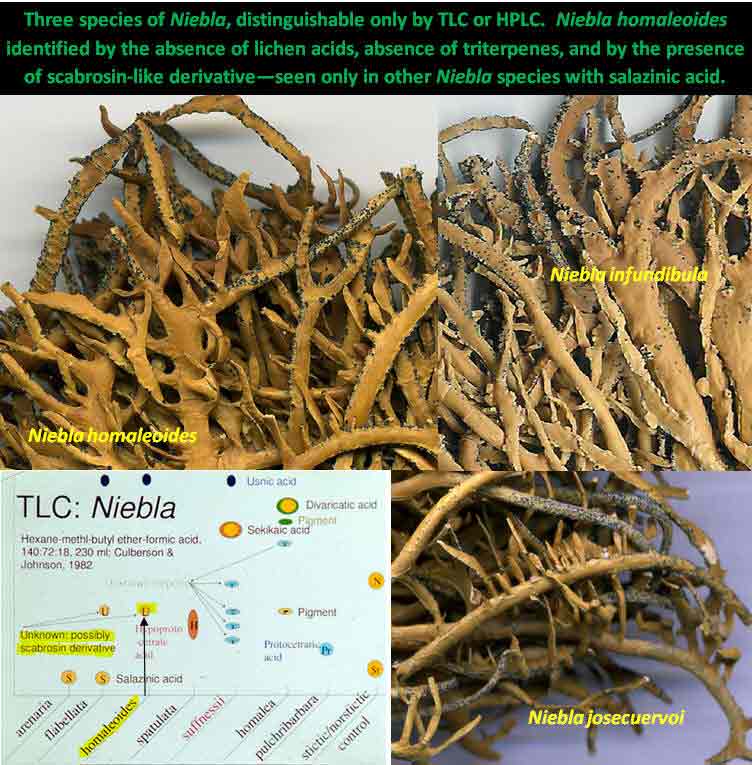
Niebla
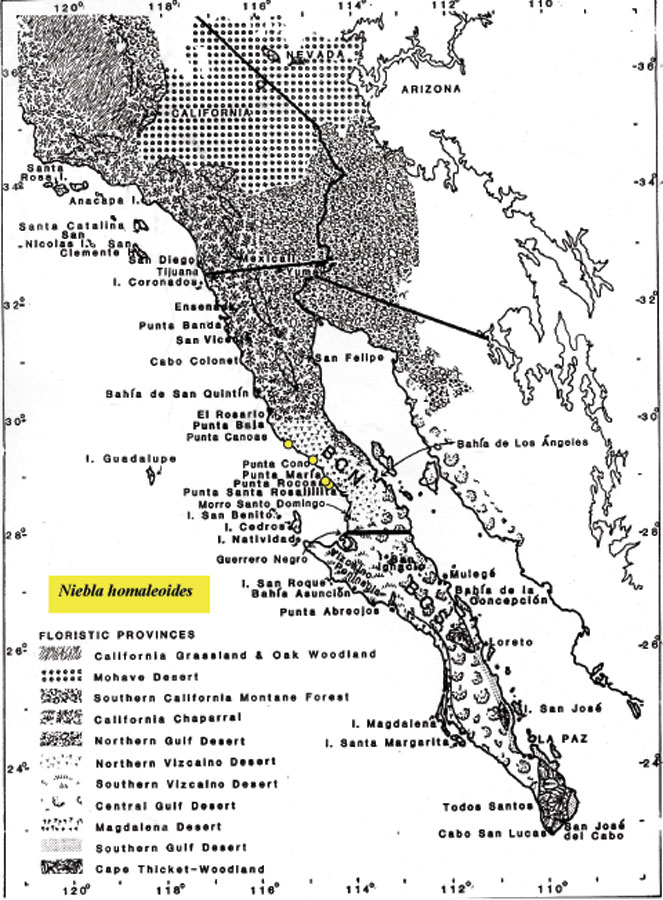
homaleoides is a rare species of lichen known only from three locations in Baja
California; two are from ridges south of Punta Negra, and a third is from a
rocky peninsula near sea level at Punta Cono. The type locality is
a large rock pile on a ridge where collected in close
association with N. josecuervoi and
N. infundibula. An
attempt was made in the field to sort these (May 1986); however, it was not
until later that it was concluded that
they could only be distinguished by their lichen substances as a result of using
thin-layer chromatography (TLC).
Their close resemblance to each other is clearly evident in the image above that
includes the TLC graphic. They are similar in overall size and branching,
have the same sublinear branch shape, have the same glossy and color
cortical features, have prominent large and abundant pycnidia on upper branches, all lack reticulate ridging, and all
show similar development in branchlets including shape and
size—somewhat elliptical and flattened—rarely seen in other
species with cylindrical branches. This seems more than coincidental considering the wide range of
morphological variation
in Niebla observed elsewhere on the ridge and generally in Baja
California (e.g., see associated species with reference to collection numbers
under N. caespitosa).
For example,
Niebla josecuervoi, which has secondary
metabolites of salazinic acid, occasionally with a scabrosin derivative and
consalazinic acid,
usually has a reticulately ridged cortex, evident in all 12 other specimens shown on
the WBA page. Did a spermatium from N. homaleoides contribute to a
spore progeny thallus of N. josecuervoi originating from an apothecium of N.
josecuervoi?
Niebla infundibula, another
relatively rare lichen known from this location, shows similar hybrid
characteristics.
Did a spermatium from N. homaleoides contribute to a spore progeny
thallus of N. infundibula originating from an apothecium of
N.
infundibula? Other specimens of N. homaleoides
from the same ridge and from Punta Cono have more flattened blades, which may be
characteristic of its true form. In the higher plant world such blending
of character traits among three species would be viewed as evidence of
introgression. Perhaps, secondary chemical substances in Niebla are allelopathic
towards related species (Molnára & Farkas 2010) in which case N.
homaleoides lacking in secondary metabolites is less competitive and more freely hybridizes?
Nonetheless, Niebla
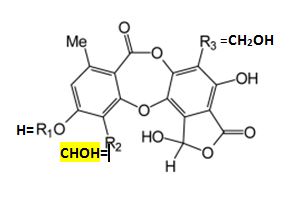
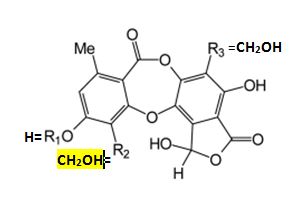
homaleoides is identified by its lack of key lichen substances.
Its chemical relationship is
clearly allied to the salazinic-acid species subgroup by the
presence of an unidentified pigment thought to be a scabrosin derivative, and further linked to the Depsidone
Species Group of Niebla by the absence of triterpenes (Spjut 1996).
This pigment has been found in N. effusa,
N. flabellata,
N. marinii, and N. josecuervoi.
Bowler and Marsh (2004) failed to account for the
chemical relationships when they
erroneously placed Niebla homaleoides in synonymy with
N. homalea (divaricatic
acid); i.e. in following their taxonomic scheme they should have lumped it with
N. josecuervoi. Just because N. homaleoides does not react
to p-phenylenediamine (PD-) does not mean
that it is related to other species that are PD-. The
resemblance to N. homalea is seen in the blade morphology in the one
specimen from Punta Cono. The relationship of N. homaleoides to the depsidone group is
also evident by the pycnidia that develop at the tips of the spine-like
branchlets, some of which are shortly bifurcate.
Chemically deficient thalli in the genus Niebla occur at one other
location, Arroyo Sauces for which a specimen is shown above. It
differs from N. homaleoides in having more flattened lacerated
branches as in
N. spatulata. This lacerated branch form was also collected at Punta Cono. Their similar
morphology at two disjunct locations would seem to represent another
acid-deficient species, referred
to as
Niebla sp. undescribed, shown above for the one from Arroyo Sauces.
The phylogeny of Niebla homaleoides might be assessed from two
acid-deficient specimens collected by Spjut and Sérusiaux from the
Vizcaíno Peninsula; however, only ITS was employed, and only one of the
two specimens showed a difference of a single mutation in the
N.
spatulata complex. Acid deficient thalli had not been reported
by Spjut (1996) from the Vizcaíno Peninsula, although he
identified four specimens in the field as N. homaleoides based on
stretched marks on the cortex, two of which confirmed by TLC.
Additional
Reference Cited:
Diaz-Allen, C., R. W. Spjut, A. Douglas Kinghorn, and H. L.
Rakotondraibe. 2021. Prioritizing natural product compounds using
1D-TOCSY. Trends in
Organic Chemistry 22: 99-114.
Molnára, K. and E. Farkas. 2010.
Current results on biological activities of lichen secondary
metabolites: A review. Z. Naturforsch. 65 c, 157 – 173.
Further References: See
Niebla.