|
Niebla and Vermilacinia (Ramalinaceae) from California and Baja
California.
|
|
|
Introduction Species Concept and Species Complexes in the Genus Niebla
Niebla homalea (divaricatic acid) species complex Niebla palmeri Species Complex Niebla spatulata species complex
How Many Species in Niebla?
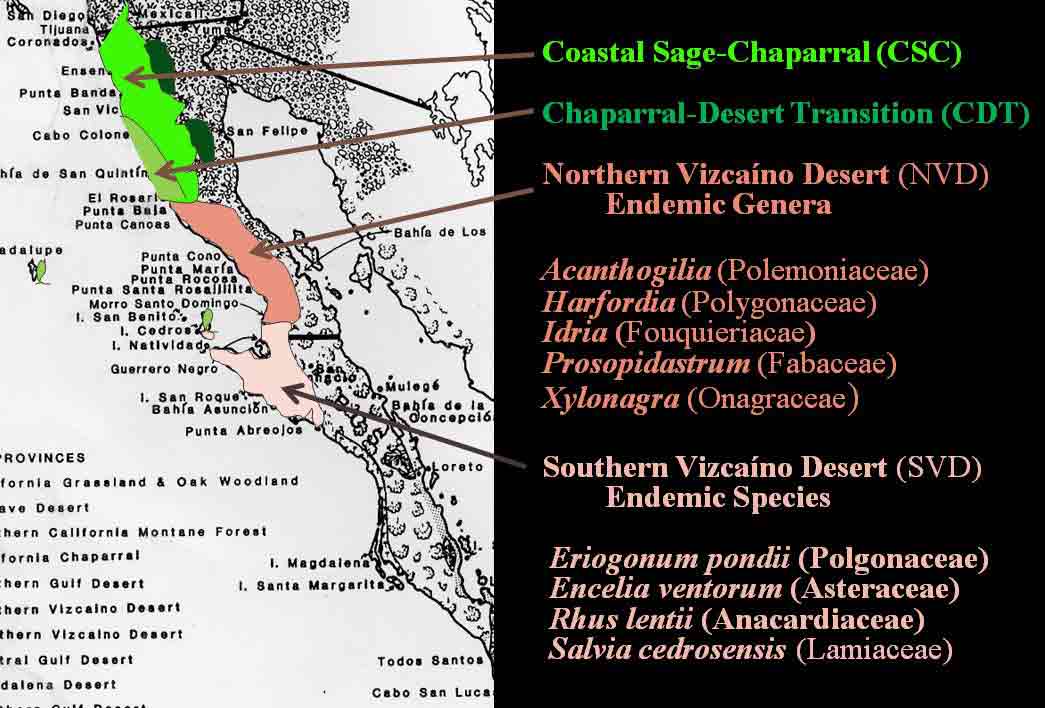
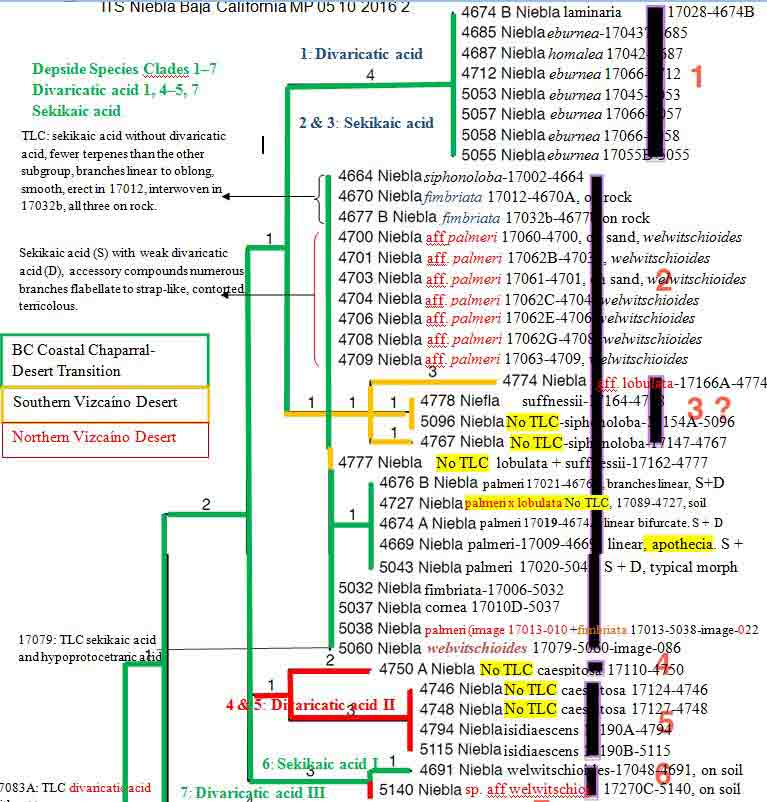
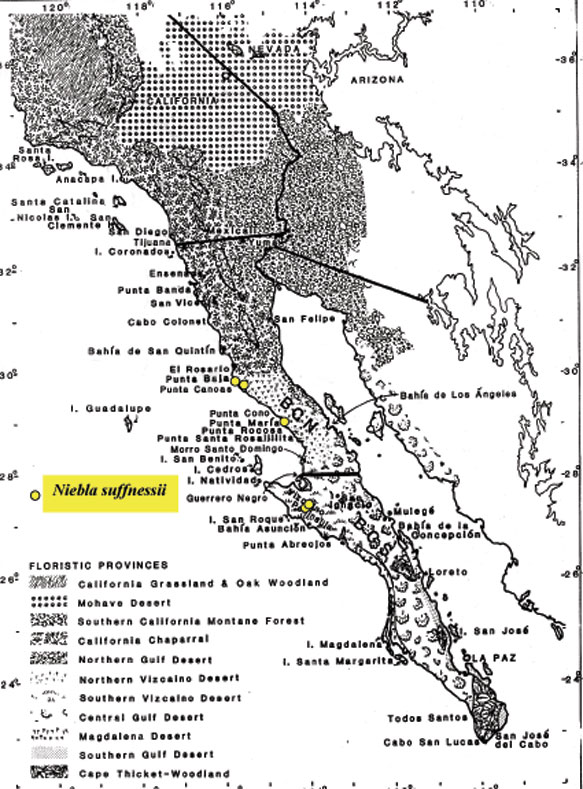
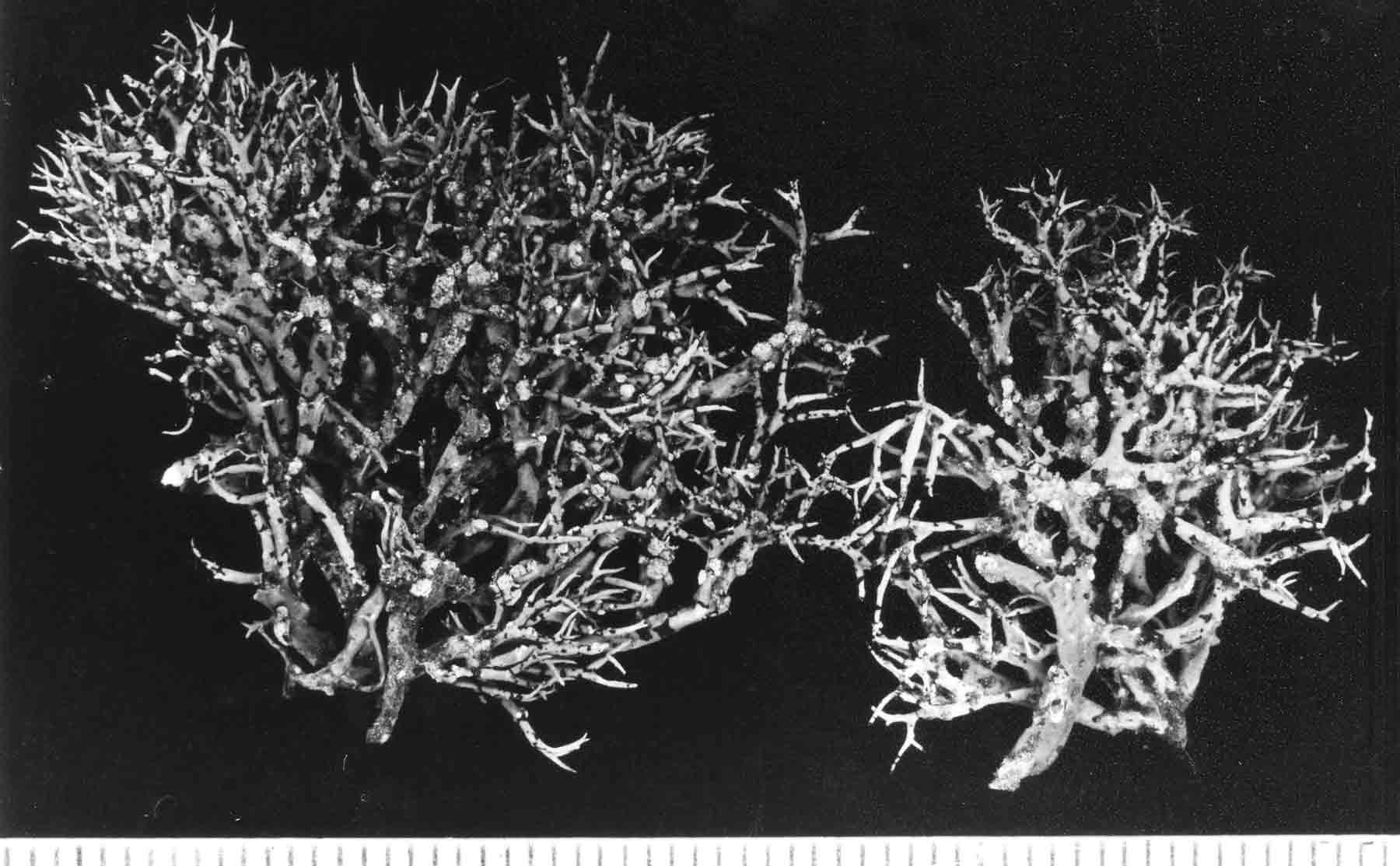
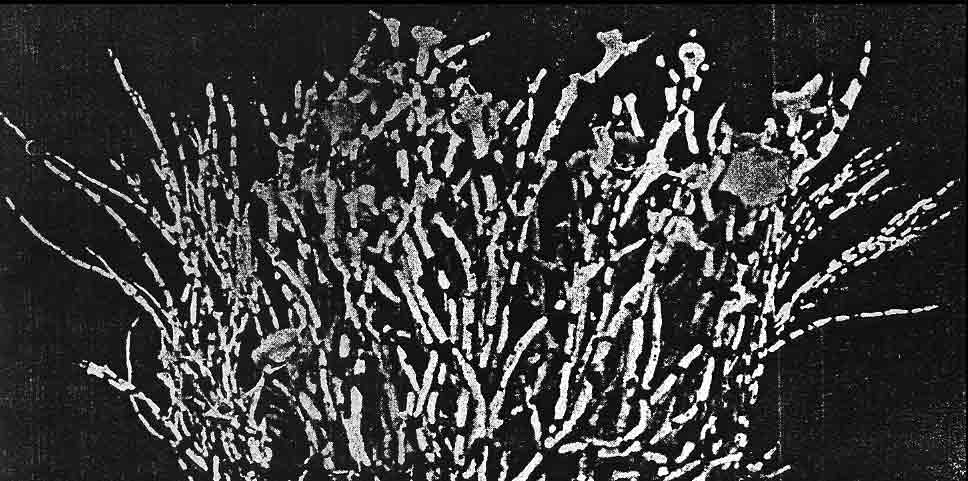
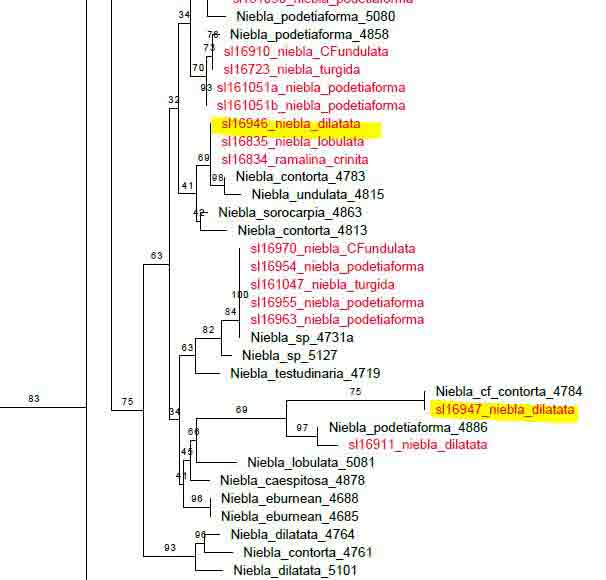
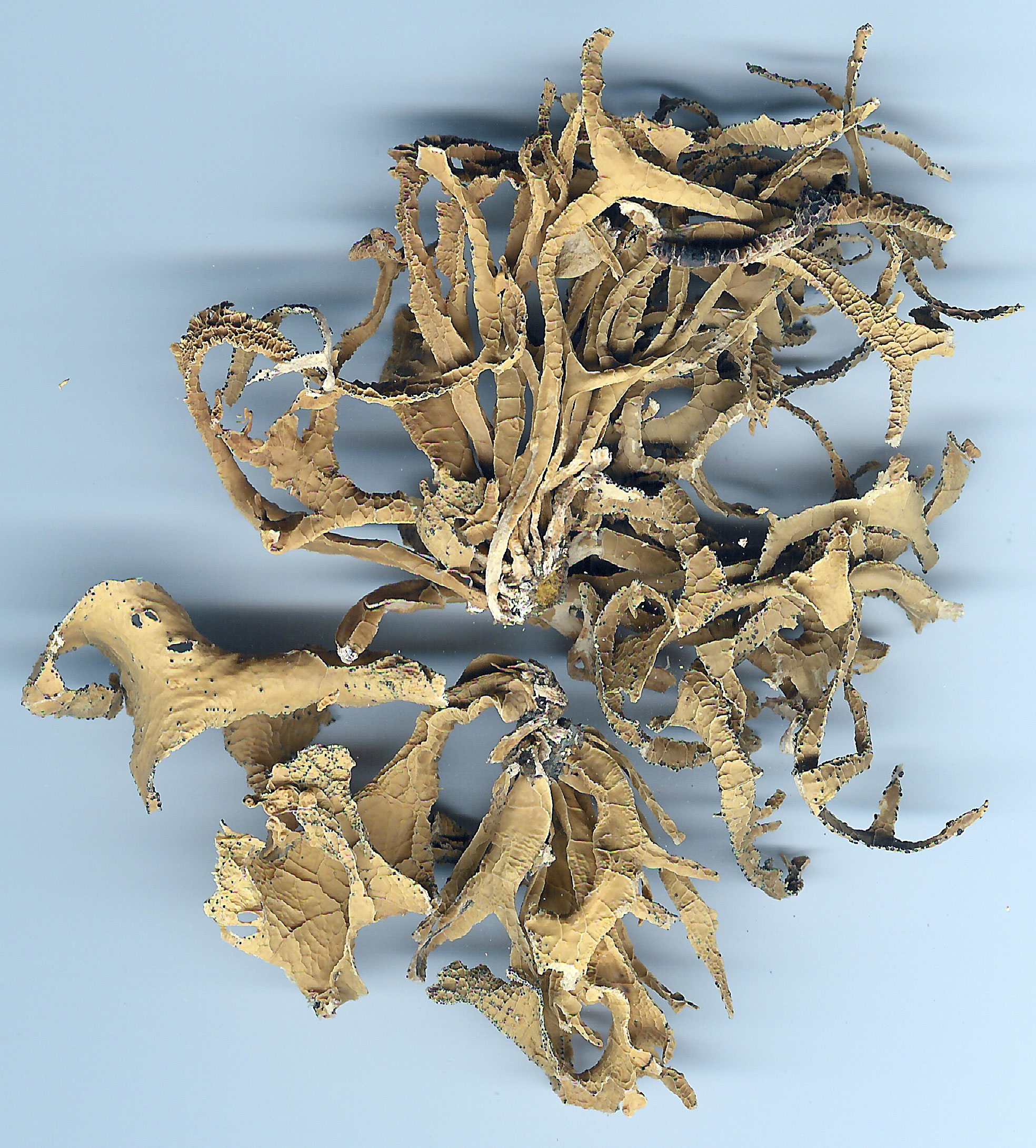
Introduction Niebla and Vermilacinia are fruticose lichens that grow along the Pacific coastal fog regions of North America, from southeastern Alaska to southern Baja California Sur, including the islands off the coast. Vermilacinia also occurs in coastal fog areas of South America from Peru to Chile (Sechura, Peruvian, and Atacama deserts), and along the coast of southwestern Africa (Namib Desert). Different types of Niebla and Vermilacinia communities are recognized based on the substrate upon which they grow (bare soil, gravel, sand, pebble, boulder). The above publication describes 70 species, 42 in Niebla (and one variety) and 28 in Vermilacinia (Spjut 1996). Five new species have since been described in Vermilacinia, four from Baja California and Baja California Sur (S�rusiaux et al. 2010; Spjut et al. 2020). The Mexico state of Baja California was previously known as Baja California Norte (BCN) as referred to in the above publication. The entire peninsular region and islands were classified into floristic provinces as shown on a Baja phytogeographic map with names for the geographic locations along the Pacific Coast where collections were made; e.g., Niebla suffnessii. Unpublished species by name (ineditus) are recognized in both genera on this website. Thin-layer chromatography was routinely employed to identify secondary metabolites in Niebla and Vermilacinia. Most species are endemic to Baja California with many narrowly restricted to the ecological regions of California and Baja California. These regions are the Coastal Sage and Chaparral (CSC), Chaparral�Desert Transition (CDT), Northern Vizca�no Desert (NVD), Southern Vizca�no Desert (SVD), and the Magdalena Region. The abbreviations for these ecoregions will be cited frequently in the text that follows.
The Magdalena Region probably has the fewest endemic species. Extrapolating from DNA phylogeny studies, these include Vermilacinia corrugata on the main peninsula where the type specimen was collected, and an apotheciate species of Niebla on Isla Santa Margarita. The southernmost occurrence of Niebla was observed (in 1996) to be isidiate species near Punta San Gregorio that are likely N. usneoides (sekikiac acid) and N. isidiaescens (divaricatic acid). Both genera are related to the � cosmopolitan Ramalina and one other genus, Namibialina, endemic to coastal southwestern Africa; all supported by DNA phylogeny (S�rusiaux et al. 2010; Spjut et al. 2020). Their phylogenetic relationship to the crustose Ramalinaceae remains unresolved. Vermilacinia was segregated from Niebla (Spjut 1995) by its secondary metabolites, predominantly terpenes, specifically the hopane triterpenoid zeorin and/or the diterpenoid [-] -16α-hydroxykaurane and by the thallus morphology of the medulla lacking chondroid strands�sclero-proso-plectenchymatous hyphae [twine-like] within more freely branched network of periclinal hyphae. The terpenes found in Vermilacinia thus far are not known in Niebla (Spjut 1996, Spjut et al. 2020; Zhang et al. 2020). It should be noted that "ceruchinol" in C. F. Culberson (1969, p. 196) is in reference to J.-M. Lehn & S. Huneck in Z. Naturforsch, 20b, 1013 (1965) where identified ��(-)-16α-hydroxykaurane, ir spectrum�; whereas in Huneck and Yoshimura (1996, p. 360) it is referred to as (-)-ent-Kauran-16α-ol), and in Rakotondraibe, Spjut & Addo (2024, p. 203, Fig. 16), it is ent-16�-hydroxykaurane. Key secondary metabolites in Vermilacinia have yet to be fully identified (Rakotondraibe, Spjut & Addo (2024); in Spjut (1996), these include "T1," "T2," etc. The terpenes in Niebla, on the other hand, are triterpenes related to stictanes and flavicanes in species formerly classified in the lichen genus Sticta, S. coronata (M�ll.Arg.), S. colensoi C. Bab., and S. flavicans Hook. f. & Taylor. The species have since been reclassified in Yarrumia (Y. coronata D.J. Galloway, Y. colensoi D.J. Galloway) and Podostictina (P. flavicans Moncada & L�cking). In Niebla, the structural differences are recognized by their nomenclatural classification of nieblastictanes and nieblaflavicanes, rare naturally occurring triterpenoids (Zhang et al. 2020). Vermilacinia is divided into two subgenera, subgenus Vermilacinia typified by saxicolous V. combeoides and subgenus Cylindricaria typified by corticolous V. corrugata; however, DNA phylogeny of Vermilacinia revealed two saxicolous species, V. combeoides and V. laevigata, in a clade sister to a corticolous clade, whereas all other Vermilacinia are saxicolous/terricolous in a sister clade (Spjut et al. 2020). DNA not obtained by Spjut et al. (2020) for other saxicolous/terricolous species such as V. acicularis, V. ceruchoides, V. robusta, and V. pumila, and related South American species�may, upon analysis of their DNA, lead to recognizing additional subgenera. Subgenus Vermilacinia includes the terricolous V. ceruchis, endemic to the coastal South American deserts; the name has a long history of being misapplied to corticolous North American species (Spjut 1996). Subgenus Cylindricaria has 11 species, four of which have been reported to occur in both North America and South America (V. cerebra, V. leonis, V. leopardina, V. tigrina). One sorediate species, V. zebrina, has a disjunct distribution between North America and Namibia. Vermilacinia corrugata (type from Baja California Sur) as circumscribed by Spjut (1996) comprises several species (V. corrugata complex) yet to be named and described, all lack ceruchinol ([-] -16 α-hydroxykaurane, Spjut et al. 2020), whereas V. tigrina and its relatives that belong to a black-banded species group, with or without ceruchinol, include V. howei, V. leopardina, V. nylanderi and V. zebrina; all need clarification (Spjut et al. 2020). An updated key to the saxicolous/terricolous species of Vermilacinia is available online at MycoKeys. 2020; 73: 1�68). The remainder of this webpage focuses on the taxonomy of the Niebla species. This includes recent studies on the phylogeny of genus and the species delimitation based on DNA (Spjut et al. 2020; Jorna et al. 2021). The general consensus is that the genus has numerous �microendemic� species, whereas Spjut (1996) recognized most phenetic species to have a broad geographical range; however, in further review here, it is suggested that microendemism in Niebla is closely associated with cryptic species complexes that include disjunct phylogeographic relationships. This may relate to northward movement of the Pacific Plate with the Baja California microplate, volcanic activity, and shifting fog regions since the mid Miocene, or earlier. Species Concept and Species Complexes in the Genus Niebla Spjut (1994) presented a paper to the American Bryological and Lichenological Society meeting titled: �What is a species of Niebla? In the abstract (American J. Botany) he stated �a broad diversity of morphotypes is evident and best classified when segregated according to the different chemotypes.� In 1996 (publication above) he referred to species of Niebla as �protean� in regard to �Proteus, an ancient Greek god who had the ability to change his shape at will.� Niebla species do appear to be �shape-shifters� (�allasomorphs�)�as first referred to on this webpage in 2005�in that a species with a distinct chemotype may vary morphologically while retaining more features in common than with other species. The term allasomorph is further applied herein to specimens representing multiple phenetic species in a DNA species clade in contrast to isomorph specimens belonging to a species that may possess specific distinctive features such as for example, sekikaic acid and isidia (e.g., N. usneoides). They are also referred to as cryptic hybrids (allasomorphs) and cryptic species (isomorphs), respectively. An example of cryptic species is seen for two specimens identified Niebla dilatata, collected by Leavitt et al., 16-946 and 16-947, in southern Baja California at Morro Santo Domingo as shown in a snip of a divaricatic acid clade (unpublished) within a phylogenetic tree. The collection numbers suggest the specimens were growing near each other. The clade shows specimens collected by Spjut and S�rusiaux (in black type) and Leavitt et al. (in red type) employed for the aforementioned publications (Spjut et al. 2020; Jorna et al. 2021). In ITS phylogenetic trees, Niebla clades often have multiple morphological species with the same chemotype as shown in the above snip. Another example is three sekikaic acid species: (1) Niebla siphonoloba�saxicolous, mostly of simple entire branches, reticulated cortex, (2) N. fimbriata�with secondary short branchlets to the main branch and intermarginal smooth cortex; and (3) N. aff. palmeri�terricolous with divaricatic acid in addition to sekikaic acid, and with branches strongly contorted and dilated with smooth or reticulated cortex. Although all regarded unresolved (Spjut et al. 2020), it is suggested here that they (allasomorphs) collectively belong to species complexes. Further, the reoccurrence of the same chemotype with the same set of morphospecies in sister clades may relate to hybridization and/or polyploidy (Campbell et al. 2015; Quattrini et. al. 2019; Ament-Vel�squez et al. 2021), or incomplete lineage sorting lagging behind clear morphological differences (Baselga et al. 2013; Myllys et. al. 2023). On the other hand, different morphotypes within the same ITS clade may actually be different species (Pedraza-Marr�n et al. 2019) as recognized for zoanthid corals (Villamor et al. 2020) whose taxonomic problems seem similar to those in Niebla except for ploidy. Also, a species complex may have multiple geographic origins (Simeone et al. 2016). The morphological classification criteria include differences in development of isidia, soredia, habit (saxicolous/terricolous), fragmentation branchlets (e.g. antler -like, comb-like), thallus size (e.g., � tufted < 6 cm, bushy), cortical ridging (e.g., ladder-like, reticulate, sinuous), and uniformity and consistency in the overall shape of branches (e.g., � linear, dilated or flattened). Major groups (or clades) of Niebla coincide with the presence/absence of terpenes; the absence associated with the occurrence of �-orcinol depsidones endemic to ecogeographical regions of Baja California. The Niebla �-orcinol depsidones usually include protocetraric acid (CDT), hypoprotocetraric acid (CDT, NVD, SVD), and salazinic acid (CDT, NVD, SVD), the latter most common, all endemic to the peninsula of Baja California and Baja California Sur extending to the southern part of Isla Cedros. The absence of triterpenes in Niebla is also phylogenetically related to the absence of depsidones (acid-deficient), a relatively rare occurrence but with frequent presence of a scabrosin derivative (A. Johnson & C. Culberson, annotation label, 1988 for specimen later selected as type for N. flabellata) in salazinic acid species. This compound may be related to 8′-O-methylsalazinic acid reported for N. aff. homaleoides (Diaz-Allen et al. 2021). The triterpenoid (depside) clade usually has either divaricatic acid or sekikaic acid chemotypes, while occasionally both may be found with one showing a stronger presence in TLC. They are widely distributed from Mendocino County in California south to Isla Santa Margarita in Baja California Sur, and west to Isla Guadalupe in Baja California. But they are not equally diverse; 21 divaricatic acid species and 12 sekikaic acid species were recognized (Spjut 1996). From a sampling of their DNA, divaricatic acid species occurred in five clades in which one included a terminal sekikaic acid clade (Spjut et al. (2020), while another sekikaic acid clade was also recognized for specimens of Niebla cornea collected in Coastal Sage and Chaparral (CSC) of California Channel Islands and in Baja California near San Quint�n (Jorna et al. 2021). Sekikaic acid specimens may also have divaricatic acid as previously indicated for N. aff. palmeri, usually weaker strength on TLC plates. Additionally, the sekikaic acid species of Niebla, which appear of relatively more recent derivation, are widely distributed with geographical diversification centered in the CDT, the Channel Islands, and adjacent (southern) California (Spjut 1996). Their center of diversity may also be viewed as the chaparral and sage regions of the Baja California microplate (Spjut et al. 2020, S1 file). This is in sharp contrast to the depsidone clade that is nearly endemic to the NVD (Spjut 1996; Spjut et al. 2020). In Baja California, as many as eight species of Niebla occur at a location (Spjut et al. 2020). Rare chemotypes in Niebla also occur. They may be viewed as hybrid exceptions to the above common pattern, often found in the CDT. A thallus may have both divaricatic and sekikaic acids (N. aff. palmeri ), or a depside with a depsidone with or without triterpenes, or sekikaic acid with either salazinic acid or protocetraric acid or hypoprotocetraric acid, or divaricatic acid may occur with salazinic acid, or may include more than one depsidone such as salazinic acid with either hypoprotocetraric acid or fumaroprotocetraric acid (SVD), or hypoprotocetraric acid with psoromic acid. These Niebla chemotypes are readily identified by TLC using solvent B' (Culberson & Johnson 1982, along with solvent G (Culberson & Johnson 1981), and running the TLC plates to the top (S�rusiaux, email comm., 2016). Solvent G, alternatively used for solvent B', does not always clearly separate divaricatic and sekikaic acids, especially if both are present and if the TLC is limited to half plate. It should be noted that Spjut identified Niebla specimens collected by Leavitt et al. (Dec 2016) for Jorna et al. (2021) based on TLC images for specimen extracts run in Solvent G, and that Spjut's TLC runs during 1986-1994 were timed to stop before the solvent reached the top of the plate. Hybridization by vegetative fusion of thalli with different chemotypes, similar to that described by Laundon (1978) for Haemotomma, is clearly evident in the Niebla spatulata complex. In filamentous Neurospora crassa, �over 150 loci have been described where mutations affect morphology (Glass, Jacobson & Shiu 2000). When a thallus is found to have both a depside and a depsidone, a DNA phylogenetic tree may show it to appear either in a depside clade or in the depsidone clade. These rare chemotypes need more study, especially where they occur among thalli growing in close proximity. For example, at one location between El Rosario and Punta Baja, hybridization is evident between N. palmeri 17077 (sekikaic acid + triterpenes) and 17080B Niebla effusa (salazinic acid) by nearby thalli identified as follows: Niebla sp. 17078-4720 with sekikaic acid, triterpenes and either hypoprotocetraric (depsidone clade) or protocetraric acid (depsidone clade), 17079-5060 N. aff. palmeri with sekikaic acid + triterpenes (Spjut et al. 2020, Fig. 7), or with addition of hypoprotocetraric acid (depside clade), and N. turgida 17080A-5062 divaricatic acid and triterpenes (typical); see N. effusa for more images. A similar hybrid species association was found near San Antonio del Mar, but DNA was not obtained for one of the specimens with the hybrid chemistry (17018). Another example is specimens collected near San Quint�n that suggest hybridization between N. effusa (17054-4695) and N. juncosa var. spinulifera (17055A-5054). A thallus containing both depsides and depsidones was confirmed by a natural product chemist based on specimens similar to N. effusa in morphology that were collected by Spjut (unpubl.) from the CDT in 1996. This phenotypic species (N. effusa) may hybridize more frequently than others in the genus.
On the other hand, these rare chemotypes could be rare species as evident in the following snip of a draft Niebla depsidone clade for a 6-locus phylogenetic tree generated by Professor S�rusiaux, Jan 2018. Collection numbers and descriptive text were yellow highlighted by Spjut to suggest hybridization; however, the thalli were collected from different locations, 17054 from near San Quint�n (SQ) shown in the previous image and 17078 shown below in a micro-clade sister to 17078 collected near Punta Baja (PB), and 17295 from Morro Santo Domingo (MD), all appeared nested within the depsidone clade instead of the sekikaic acid clade, or in a separate monophyletic sister clade. TLC showed both salazinic acid and sekikaic acid accompanied by triterpenes for all three specimens, including 4872, also with salazinic acid. Additionally, 17078 showed protocetraric acid with triterpenes in another TLC run.
Morphological evidence for hybridization includes distinct morphotypes that occur together but differ in their chemotype. An example is thalli on Mesa Camacho, Niebla angulata Spjut ineditus (salazinic acid) and N. sinuata Spjut ineditus (sekikaic acid). Both share unusually raised sinuous cortical ridges. Less prominent sinuous cortical ridges are generally seen in sekikaic acid species (Spjut 1996), thus, one may conclude that this feature evolved with N. sinuata on Mesa Camacho where subsequently N. marinii (salazinic acid, typically with smooth cortex) acquired this trait through hybridization, thus, N. angulata evolved. Additionally, DNA phylogenomic data reveal many genealogical species complexes. These include indistinguishable phenotypic specimens (isomorphs) with different genotypes (cryptic species) that occur in close proximity and/or at disjunct locations such as in the Niebla homalea species complex (Spjut et al. 2020, Fig. 7). This author refers to them as cryptic hybrids when they differ morphologically (allasomorphs), or they may in fact be different species as suggested earlier, considered unresolved by Spjut et al. 2020 and Jorna et al. 2021, Authorea. These DNA studies focused on comparing phylogenomic data to the taxonomy of Spjut (1996), evaluating endemism by number of different localities where collected, and validating �species boundaries� by employing various statistical analytical methods on single locus and concatenated mult-locus sequences to delimit species. It may be noted that a gene sequence may be on average more distant from another sequence of the same species than from a sequence randomly sampled from another species (Jiao and Yang, 2021). Jorna et al. (2021) employed a reference genome of N. homalea collected from Shell Beach in Sonoma County (Ametrano et al. in Duong et al. 2021) for restriction site-associated DNA sequencing (RADseq) to generate > 25,000 loci from which sets of the most variable loci were selected, specifically, or randomly, and employed various in-depth statistical analyses, but �were unable to infer robust, consistent species delimitations.� A Niebla species complex may be closely associated with several or more paraphyletic or polyphyletic species complexes shown in the following image. Collectively, a reticulate pattern of speciation seems evident (e.g., Persson et al. 2020). These may result from incomplete lineage sorting , chromosomal rearrangement, horizontal gene transfer, transposable elements, dikaryosis, or multiploidy compositions (Tripp et al. 2017; Pizarro et al. 2019, Grande et al. 2021; Ament-Vel�squez et al. 2021).
Examples of species complexes in Niebla are described in further detail as follows. Niebla homalea (divaricatic acid) species complex Niebla homalea was circumsribed by Spjut (1996) to have a thallus with divaricatic acid and divided into sublinear � segmented (transversely ridged and/or cracked) branches. He interpreted this species to be widely distributed from northern California to northern Baja California (Spjut 1996); however, DNA phylogeny (Spjut et al. 2020) reveal this to be not exclusively monophyletic. When the polyphyletic clades are viewed geographically, cryptic species and cryptic hybrids (unresolved species) are evident in both California and Baja California, and may include allasomorphs such as N. eburnea and N. testudinaria, that might prove to be a genealogical distinct species distinguished by isodivaricatic acid instead of divaricatic acid, at least for the type locality of N. eburnea from near Colonet, Baja California.
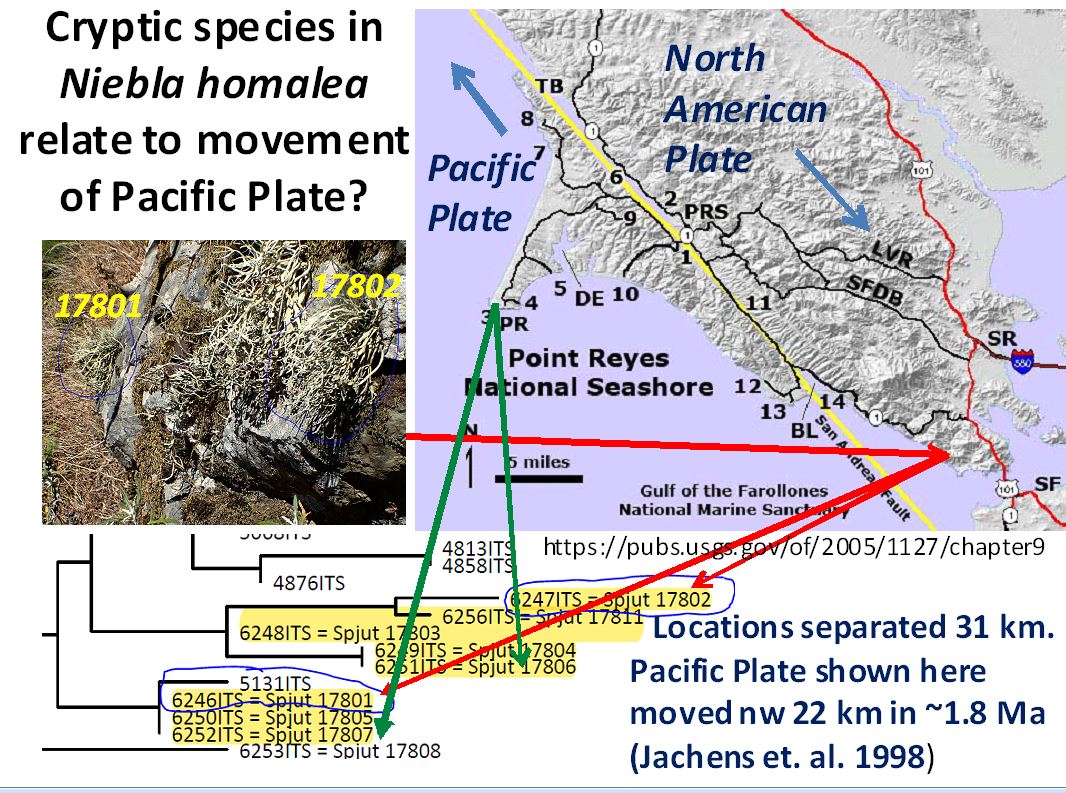
The above image shows ITS phylogeographic relationships of the Niebla homalea species complex for specimens collected in Marin County south of Stinson Beach, 17801 to 17805 (North American Plate), and at Point Reyes�17806- 07 (Pacific Plate). This complex includes specimen 17811 from the Monterey Peninsula (Pacific Plate, outside map); specimens 17805 and 17811 were identified N. eburnea, all others N. homalea; 17801 and 17802 are labeled in the photo showing where they grew on a large rock before collected, and a red arrow leads to their location on the map, and then to their position in the relevant portion of a phylogenetic tree. These latter two thalli (17801, 17802) occurred in two ITS clades. So also does 17806 and 17807 occur in different clades, collected near each other at Point Reyes (Pacific Plate) indicated by the green arrows. Niebla eburnea 17811 from the Monterey Peninsula on the Pacific Plate�not phylogenetically resolved in view of 17805 from near Stinson Beach (NA Plate) in another clade�is shown in a subclade sister to N. homalea 17802 collected at Pt. Reyes (Pacific Plate).
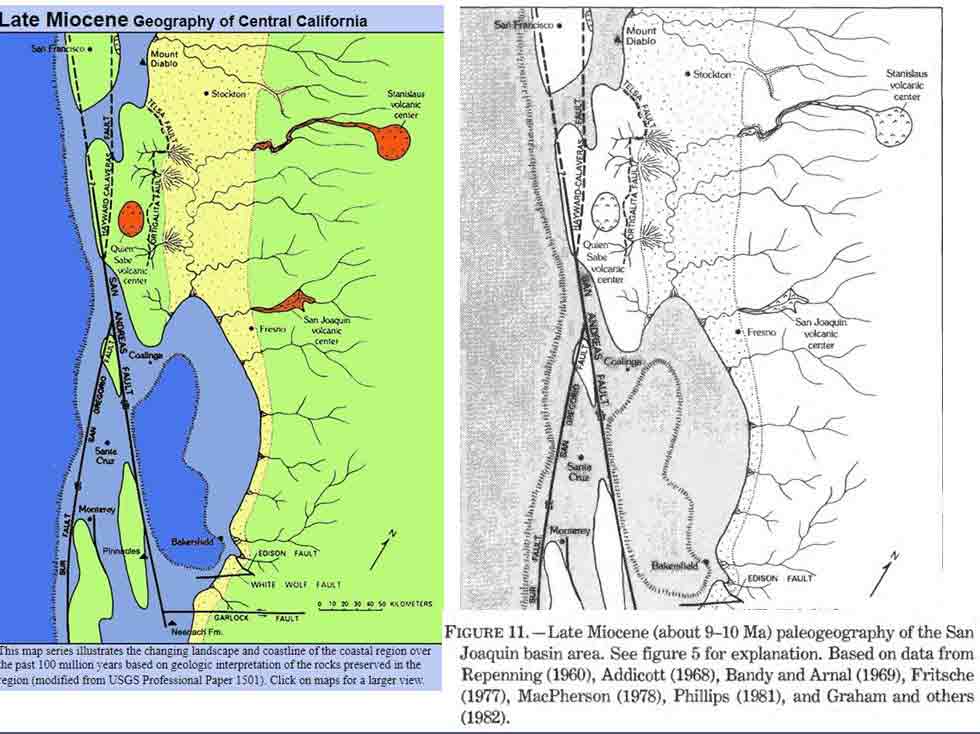
Although cryptic species in the N. homalea species complex may relate to isolation of ancestral thalli by movement of the Pacific Plate northwest relative to that of the North American plate during the past 1.8 million years (Spjut et al. 2020)�as indicated on the preceding map of the Point Reyes Peninsula, that of N. eburnea on the Monterey Peninsula appears older. The Monterey rocks, which are of the same type and age as that at Point Reyes, are estimated to have been displaced 180 km in 5.3 million years (McIlrath 2010). An older N. eburnea is supported by the longer branch in the phylogenetic tree, sister to N. homalea at locations where collected on the Pacific and North American plates. So it would seem that N. eburnea has changed little in the past five million years, except for its cryptic traits (Spjut 2022, cited in Rakotondraibe HLR, Spjut RW, Addo EM. 2024). The Monterey Peninsula perhaps was an island where N. homalea once thrived isolated from an older N. homalea/N. eburnea complex sister to that in Baja California, while N. homalea (excluding the allasomorph N. eburnea) is mostly absent from southern California including the Channel Islands. The following maps show how the former California coast might have been during the Late Miocene (9�10 mya), the color rendition is from Geology Cafe website, the original gray created by J. A. Bartow (1991): The evolution of the San Joaquin Valley, USGS paper 1501. Observe former islands off the coast of California, and that Monterey was positioned at the latitude of Bakersfield, California. One might expect that such islands may have existed off Baja California before it displaced in late Miocene from mainland Mexico near Puerto Vallarta.
The putative type locality for N. homalea is Point Reyes in Marin County, California (Zhang et al. 2020). Additionally, historical collections from Point Reyes include Niebla disrupta, distinguished from N. homalea by having sekikaic acid (Spjut 1996). Since N. disrupta can be distinguished by TLC, it is not considered a cryptic species with respect to N. homalea, nor is it likely to be closely related. Although N. disrupta was not identified among samples collected at Pt. Reyes for studies by Spjut et al. (2020) and Zhang et al. (2020), it may be expected to belong to a sekikaic acid species complex such as that described below for Niebla fimbriata and Niebla palmeri. The phylogeographic relationships for specimens identified Niebla homalea and N. eburnea at disjunct locations�between San Quint�n (Baja California) and Marin County (California)�is interesting. In the intervening area�southern California, including the Channel Islands�is N. cornea that differs from N. eburnea by producing sekikaic acid instead of divaricatic acid. DNA phylogeny for specimens of N. cornea collected from San Clemente and Santa Barbara islands and from near San Quint�n in Baja California by Leavitt et al. (Jorna et al. 2021) support the Spjut (1996) taxonomy of this species essentially for its entire geographical range absent data from the type locality at Morro Bay, California where probably extirpated. The uplift of the Channel Islands is the result of the northwest movement of the Baja California plate�captured by the Pacific Plate�plowing into California, beginning ~5 mya (Atwater 1998). Thus, the N. homalea species complex was possibly fragmented by uplift of the Channel Islands and Transverse Ranges�and taking into consideration the phylogeography of the N cornea clade�the N. homalea species complex would appear older. Species radiation within Niebla is estimated to have begun ~ 13 mya (Spjut et al. 2020).
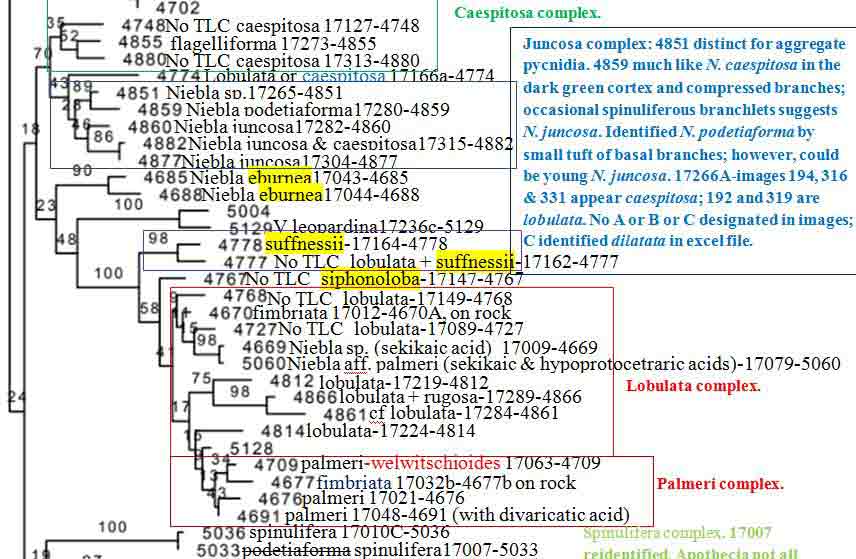
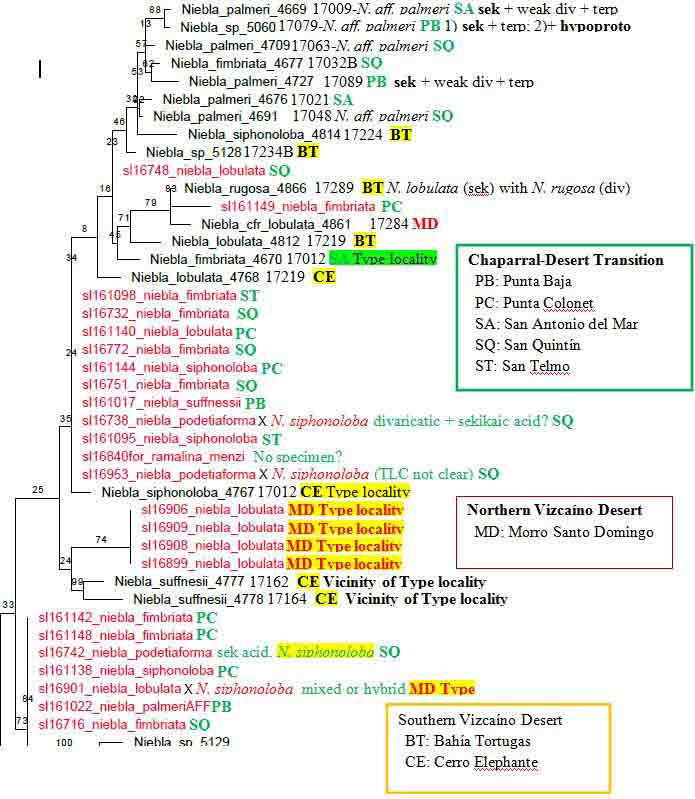
Niebla fimbriata and N. palmeri (sekikaic acid) species complexes. Niebla fimbriata Species Complex Typical Niebla fimbriata is saxicolous characterized by primary branches with a relatively smooth cortex between undulating branch margins from which arise shorter secondary � pinnately arranged branchlets that break off. The type is from a lava mesa above (east of) San Antonio del Mar (SA) where specimen 17012-4670A shown above was collected. The DNA supports the species as described by Spjut (1996) to include a specimen collected at Bah�a de San Quint�n (SQ), 17032b-4677, and another cited in (Jorna et al. 2021, Authorea) collected ~ 10 km south of the type locality near San Telmo (sl-161098). Both locations include additional sekikaic acid species in the N. fimbriata species complex identified N. siphonoloba and N. aff. palmeri (allasomorphs), and N. lobulata in one of the two sister subclades, which a clade sister to a N. lobulata clade is represented by two specimens from the type locality (Morro Santo Domingo) and another from the SVD, the clade described later in further detail. However, a draft ITS phylogeny of (17-) 18 (-19, color coded for geography by Spjut) Niebla species groups revealed N. fimbriata to include saxicolous N. siphonoloba collected on lava at SA and terricolous N. aff. palmeri (N. welwitschioides ineditus)�collected on beach sand near the shore of Bah�a Falsa, Bah�a de San Quint�n (SQ), shown in the above image, represented by 17062, compared more closely to typical N. palmeri (17021) growing on rocky soil near SA . Niebla siphonoloba is phenotypically distinguished by sparingly branched basal branches with reticulate-transversely oriented-cortical ridges in sharp contrast to the welwitschioid N. aff. palmeri divided into broad strap-shape contorted branches bearing resemblance to the gymnosperm genus Welwitschia, and by having divaricatic acid in addition to sekikaic acid. It may be noted that N. fimbriata has only sekikaic acid at the type locality (SA), where at SQ and further south along road to Punta Baja it may also include trace of divaricatic acid.
Niebla palmeri species complex The Niebla palmeri species complex is recognized by terricolous thalli having sekikaic acid and triterpenes. It overlaps with the N. fimbriata complex by the phylogenomic data and hybrid chemotypes, which may include depsidones. A sampling of a local population of 10 thalli on a sand dune at SQ (17060 to 17063 & subdivided)�all collected in close proximity (� within several meters of each other)�included seven identified Niebla aff. palmeri (N. welwitschioides ineditus ) that were consistent in their position in phylogenetic trees, morphology, and chemistry, while the remaining three were of two phenotypic salazinic-acid species (N. limicola, N. arenaria). However, also collected on sand a short distance to the north (< 1 km) was N. aff. palmeri (N. welwitschioides), 17048-4691, that differed by three mutations, and sister to a specimen collected at Morro Santo Domingo (MD), 17270c-5140, identified N. aff. welwitschioides (ineditus). The geographical distance between 17048 (SQ) and 17270c (MD) is ~ 375 km (Google Earth). Also, a specimen collected along the road from El Rosario to Punta Baja (PB) identified N. aff. palmeri (N. welwitschioides) was in the same clade as the 17060-63 specimens from SQ but found with hypoprotocetraric acid in addition to sekikaic acid and triterpenes in one of two TLC runs (or two thalli), the other with only sekikaic acid and triterpenes. Additionally, this diverse phenotypic clade included a specimen collected from volcanic rocks on a mesa above Punta Baja, 48 km south of SQ (17079-5060) where it occurred in close association with other thalli containing protocetraric acid or hypoprotocetraric acid in addition to sekikaic acid; the type specimen for N. pulchribarbara near SQ contains only protocetraric acid (Rundel & Bowler in Rundel et al. 1972), which was upheld as a distinct species by Spjut (1996) but not supported in DNA phylogeny from a specimen collected at SA (Spjut et al. 2020). Typical Niebla palmeri (type from Islas de los Coronados) has only sekikaic acid (with triterpenes) and relatively narrow sublinear branches, similar to 17021 on paper bag shown above (Spjut 1996; Spjut et al. 2020, Fig. 7-extract #4671). DNA from four of five phenotypically similar specimens collected on a lava mesa above San Antonio del Mar (SA), ~ 82 km north of SQ, were in a separate ITS subclade (or subgroup) from the welwitschioid N. aff. palmeri. In the 6-locus phylogenetic tree of Spjut et al. (2020, Fig. 7), the SA specimens in a subclade are sister to a N. lobulata subclade in which two of three specimens were collected from Morro Santo Domingo (MD), the type locality; the third N. lobulata specimen was collected in the SVD northwest of Bah�a Tortugas (BT). Thus, the N. fimbriata and N. palmeri species complexes would seem to include many phenotypic species, appearing to have both cryptic species (isomorphs) and cryptic hybrids (allasomorphs) identified N. palmeri and N. siphonoloba. This variation may relate to rapid speciation and incomplete lineage sorting of ancestral lineages (Parins-Fukuchi et al. 2021), which may involve hybridization.Disjunct phylogeographic patterns are seen in other sekikaic acid species clades such as with the polyphyletic N. lobulata (Spjut et al. 2020, Fig. 7). As mentioned above, this species is actually supported as shown in a 6-locus phylogenetic tree by three specimens, two from the type locality, Morro Santo Domingo (MD, 7 17284-4861 and 17289-4866), and the third from the SVD (Vizca�no Peninsula) northwest of Bah�a Tortugas (BT, 17219-4812). Niebla lobulata is also similarly supported from the type locality by four specimens collected by Leavitt et al. shown in a draft ITS subclade sister to N. suffnessii (unpubl. phylogenetic tree generated with DNA sequences from Spjut et al. 2020 and Jorna et al. 2021); the two subclades are sister to a large clade of specimens collected mostly in the CDT; the exceptions, which appear basal in the successive smaller clades, were collected in the SVD near Cerro Elefante (CE, N. siphonoloba, near type locality) and BT. For example, near BT specimen 17219 was 17224-4814 identified N. lobulata appearing basal in a clade to specimens from three locations in the CDT, one (1) collected from Punta Baja, (2) two collected from SQ and (3) two collected from SA (Spjut et al. 17020-5043 and 17021-4676 in Fig. 7). But none were collected in the NVD other than at the type locality, MD. The absence of collections from the NVD is a pattern also noted for related species within genera of angiosperms (Spjut et la. 2020, Suppl. data file 1). One may also conclude that N. lobulata, N. siphonoloba and N. suffnessii are microendemic species that have hybridized with N. fimbriata and N. palmeri in the CDT as a result of the Pacific Plate drifting northwest into California during the past six million years. Niebla spatulata (hypoprotocetraric acid/salazinic acid) species complex
Disjunct phylogeographic patterns are also evident in the depsidone clade, presented above for the Niebla spatulata species complex extracted from an ITS phylogenetic tree generated by Professor S�rusiaux, color coded by Spjut to show phylogeographic relationships. This species complex is nearly endemic to the SVD, usually as localized populations in which the individual thalli occur in close proximity to each other. The N. spatulata thallus has basal branches terminally flattened and crinkly in various tortilla-chip like shapes with conspicuous pycnidia appearing larger than in most other species in the genus. Individuals within the population usually contain salazinic acid and/or hypoprotocetraric acid with 4-O-demethylnotatic acid as major or minor component associated conhypoprotocetraric acid, occasionally with no detectable lichen substances (acid deficient, N. aff. homaleoides). Among 29 specimens sampled in 2016 from three locations on the Vizca�no Peninsula northwest of Bah�a Tortugas (BT), was 17183A-4786) that was more phylogenetically related to specimens (Species Group 10) collected ~ 300 km further north, near Punta Baja (PB, e.g., 17080B-5062), than to the specimens growing nearby within centimeters of each other, e.g., 17184B-4788, whereas other specimens growing nearby the BT specimen, e.g., 17186B-5111, belonged to a third ITS clade sister to 17183A-4788. All three aforementioned specimens were identified N. flabellata based on having salazinic acid instead of hypoprotocetraric acid (N. spatulata), but the latter chemotype was usually present in close proximity, appearing in all three or four subclades with N. flabellata (Spjut et al. 2020, Fig. 7). Since the chemical differences were not discriminated by ITS nor by the 6-locus dataset, that also included two acid-deficient specimens of N. homaleoides except for one mutation within the large depsidone clade, the three ITS lineages represent three cryptic species within the Niebla spatulata complex. These phylogeographic relationships are not readily apparent in Spjut et al. (2020, Fig. 7) because political boundaries were applied (CDT not distinguished from NVD) to the phylogenies (trees) that were generated by extract numbers without the associated collection number, which their data are in a separate supplemental file. Notwithstanding, the three subclades of the N. spatulata species complex are clearly depicted in the 6-locus phylogenetic tree of Spjut et al. 2020, (Fig. 7) that was determined to be only one species (N. spatulata) by statistical analyses of the SVD clade, although two lineages could be recognized; however, a second (N. flabellata, 17183B-4786) is sister to the large NVD depsidone clade with exception to one specimen shown monophyletic collected near Guerrero Negro (N. limicola, 17130-4751). This lends support to three cryptic species in the N. spatulata. Additionally, in Fig. 7 of Spjut et al. (2020), Niebla limicola, 17130-4751, collected on sand dunes southwest of Guerrero Negro, appears phylogenetically distinct�at the type locality. Occurring with N. limicola was a distinct morphotype with a distinct chemotype in having both hypoprotocetraric acid and psoromic acid. Identified Niebla sp. (17132-4752), its phylogeny was affiliated with unresolved depsidone species collected in the CDT near El Rosario and San Quint�n. As previously noted, this phylogeographic relationship is also seen in the depside clade for both divaricatic and sekikaic acid subclades (Spjut et al., Fig. 7, sekikaic acid N. lobulata, N. suffnessii). Similar disjunctions in vascular plant species were mentioned by Spjut et al. (2020, Suppl. Data file S1). They suggest that the NVD was perhaps once an island separated from Baja California, most likely before the peninsula separated from mainland Mexico. Niebla flabellata may also be recognized with DNA support, but not as circumscribed by Spjut (1996), possibly from only one of three specimens collected near the type locality, ~4 km south of Puerto San Andr�s�where found associated with the same divaricatic acid species reported at the type locality, N. flagelliforma, N. caespitosa (Spjut 1996, Spjut et al. 2020, extract 4883). It�s phylogeny was shown monophyletic (96%) basal to most depsidone species (Spjut et al. 2020, Fig. 7). Spjut and S�rusiaux visited the type locality for N. flabellata in 2016, but only collected the associated N. caespitosa (divaricatic acid). It is also interesting that phylogenomic ITS differences in the coastal fog lichen Dendrographa leucophaea (Roccellaceae ) parallel phenetic differences in isomorph specimens of Niebla laminaria (divaricatic acid) at Punta Banda and on San Nicolas Island. Lohtander et al. (1998) reported that the most closely related phylogenomic specimen was not necessarily from the thallus growing nearby, but one at a more remote location, e.g., Punta Banda in Baja California and San Nicolas Island in California. This study focused on whether asexual reproduction (via fragmentation) vs. sexual (via spores) represented different species based on samples from the nine widely separate locations: Channel Islands (San Nicolas, Santa Catarina, Santa Cruz), Diablo Canyon in San Luis Obispo County, and Punta Banda, San Quint�n, San Juanico, and Isla Guadalupe in Baja California and Baja California Sur. The results yielded a monophyletic group divided into four clades, two mixed and two segregated into separate sexual and asexual clades, but no formal taxonomic distinction made. Also, no clades were specific to a geographical location; however, the largest clade was comprised of three specimens from near San Quint�n and two from San Nicolas Island, whereas all other specimens in the remaining three clades were from more distant locations, e.g., �Clade 4� (100% support) included a subclade with 63% support comprised of two specimens, one from San Juanico in Baja California Sur, the other from San Nicolas Island; they are separated by a distance of 1,030 km (Google Earth). Unresolved Niebla pulchribarbara Complex The Niebla pulchribarbara complex includes unresolved protocetraric acid N. pulchribarbara, hypoprotocetraric acid N. brachyura, acid deficient N. homaleoides, and salazinic acid N. josecuervoi, N. arenaria, N. effusa, and N. marinii. All belong to the nonterpenoid (depsidone) clade. Excluded are N. flabellata (salazinic acid), N. limicola, and N. spatulata (hypoprotocetraric acid) because the species appear supported at or near their type locality (N. flabellata, N. limicola, N. spatulata) or belong to a well-defined species complex (N. spatulata). Two of the species were named after field assistants, Richard Marin who accompanied Spjut on many expeditions to Mexico BC and BCS and Jose Cuervo who accompanied Rundel et al. (1972) in their study of Niebla (�Desmazieria�) at Bah�a de San Quint�n. Jose Cuervo is also a well-known brand of tequila, named after a family-owned tequila company born in Mexico 1795. Except N. homaleoides, all were treated as synonyms of N. josecuervoi by Bowler and Marsh (2004). Niebla pulchribarbara (protocetraric acid), N. brachyura and N. homaleoides (hypoprotocetraric acid) were distinguished from other depsidone species by their chemistry, whereas the remaining salazinic acid species were distinguished by morphology.
The Morro Santo Domingo Niebla brachyura (Spjut 1996) included two of four specimens representing three species (BPP ) with this chemotype, all in separate clades or subclades (Spjut et al. 2020, Suppl. S3, S5, Fig. 7). They possibly can be distinguished by the terminal antler-like branching of the thallus that might be compared to antlers of moose�tall curved, low wide spreading, dilated in part, 17262-4848), or elk (tall sharp, unequally bifurcate, 10270a- 4852), or reindeer-like (sharp pointed, � equally bifurcate, N. arenaria), although antler-like branching is highly variable in the more common phenotypic N. arenaria (salazinic acid). However, 17262 is unusual in having abundant pycnidia immersed below the cortex; otherwise, most specimens of N. brachyura have pycnidia visible at apex and below on cortical ridges of terminal branchlets (Spjut 1996). Unfortunately, N. brachyura was not collected by Spjut and S�rusiaux at the type locality near Puerto San Andr�s, where in 1985 it was very rare, known only from a single specimen pulled out of a paper lunch-size bag containing a mixed sample of many thalli of N. flabellata (salazinic acid, type locality) and N. caespitosa (divaricatic acid). A third specimen, 17082-5063�collected in the southern region of the CDT near Punta Baja�has a thallus similar to N. effusa; indeed, phenotypic N. effusa was nearby (17080B-5062). The thallus was mat-like with horizontal spreading (not upright) branches, with cortex strongly reticulately-ridged, with branching at right angles, and with equal terminally divided long recurved branchlets. As noted earlier under the N. fimbriata and N. palmeri species complexes, hypoprotocetraric acid was also found in a thallus with sekikaic acid at the same location, while a thallus with protocetraric acid and sekikaic acid (17078-4720) was also nearby. One might conclude that terricolous N. effusa and N. palmeri hybridize more frequently with other species of Niebla. As described above, specimens that differ in both morphology and chemistry may occur within a clade whether collected from the same location or from different locations (Spjut et al. 2020; Jorna et. al,). Although the Niebla DNA studies have been for the most part unable to correlate phylogenomic species with morphological characters, these phenotypic differences in many cases are difficult to ignore, and perhaps should be applied in future phylogeny studies of Niebla (DeLong-Duhon & Bagley 2020; L�cking, Leavitt & Hawksworth 2021) . Estimates on the number of DNA species in Niebla analyzed by various statistical methods applied to multi locus sequences vary widely. In the (Spjut et al. 2020) study in which 29 phenotypic species (including one sp.) were identified according to Spjut (1996), the ABGD bar code method that recognized (17-) 18 (-19) species groups�upon further statistical analysis (Puillandre et al. 2012, 2021) yielded 25 species, 22 in the depside (terpenoid) clade and three in the depsidone (nonterpenoid) clade, whereas the BPP (Bayesian Posterior Probability) analysis�combining six-loci�recognized 33 species, 24 in the depside clade and nine in the depsidone clade. It is interesting that the BPP analysis gave three times more species for the depsidone clade compared to just two additional species for the depside clade. Jorna et al. (2021) delimited a statistical median range of 17 species from 40 specimens collected by Leavitt et al. in 2016�representing 22 phenotypic species and one variety according to Spjut (1996) taxonomy as presented in phylogenetic trees�based on Assemble Species by Automatic Partitioning (ASAP; Puillandre et al., 2012, 2021) of the ITS locus. They analyzed the sequences by genomic species delimitation under the multispecies coalescent model using BPP/Genealogical Divergence Index (GDI) for subsets of the most informative and variable loci from a concatenated RADseq loci dataset of 25,086 loci. However, �a 29 species model� could also be recognized. Three of five phenotypic species identified in the depsidone clade were recognized by statistical analyses. However, the lower species number, of median range, 17 species, may be difficult to reconcile in view of the fact that eight species may occur at one location. In view of the preceding species complexes that generally show two to three times more cryptic species than the phenotypic (chemo-morphology) method of Spjut (1996), the number of Niebla species is likely to be closer to the higher estimates given for the DNA methodical species analyses such as measured by BPP genealogical divergence index (gdi) scores. For example, the N. spatulata complex includes three species that usually occur in close proximity while appearing distinguishable only as a species complex by thallus morphology, chemistry, and geography. All three cryptic species are endemic to the SVD. Thus, micro-endemism in this case is closely associated with cryptic species. Moreover, sampling of the entire Niebla population was rather limited based on very broad species concepts of Spjut (1996). In view of the proposed high number of Niebla species that probably exist, the 101 specimens in Spjut et al. (2020) and 40 of 182 specimens in Jorna et al. (2021) is only a mere sampling of the 42 phenotypic species in Spjut (1996) whose study included specimens from northern California, the Channel Islands, Isla Cedros, Isla Guadalupe, and other minor islands. With exception to N. cornea (sekikaic acid) from San Clemente and Santa Barbara islands (Jorna et al. 2021), relatively few specimens for each of the probable existing species of Niebla were represented in the DNA phylogenies. One might also expect that some of the disjunct occurrences reported above may appear intermittent with larger sample sizes, especially if collected from mesas and ridges in the NVD where many of the types were collected but not visited by the collectors. Speciation in Niebla is suggested to have largely occurred during the advance and retreat of glaciers since the mid Miocene (Spjut et al. 2020). As fog became localized from reduced upwelling during glacial periods (Herbert et al. 2001; Johnstone & Dawson 2010, Ganeshram et al. 2021 ), Niebla populations likely fragmented leading to allopatric speciation. When Niebla populations expanded their range during shorter interglacial cycles, they may have come into contact and hybridized. �Climate driven vicariance� has been suggested for species complexes in other organisms, e.g. aquatic vertebrates (Hawlitschek et al. 2012; amphibians (Thom� et al. 2020) that can involve historical hybridization as reported for the angiosperm Gentian family (Fu et al. 2021) and the lichen Rhizoplaca melanophthalma group in subalpine habitats (Keuler et al. 2020, 2022). Hybridization was also suggested for Niebla (Spjut 1996) with reference to studies in the genus Cladonia by C. Culberson et al. (1988) 13 genotypes could be found in a mat of C. chlorophaea associated with variable insertions, mostly Group I introns rDNA (DePriest 1994). Hybridization by fusion of thalli is evident in the N. spatulata complex where they happen to make physical contact, which may involve four chemotypes from two or more species (N. contorta, N. homaleoides, N. spatulata). Current change in climate appears related to less fog occurrence and duration as a result of increasing urbanization (Williams et al. 2015). Population growth in coastal areas in the municipality of San Quint�n, for example, increased from 4,777 in 2010 (Wikipedia) to 117,568 in 2020 (Wikipedia). However, the disjunct geographical relationships within species complexes also indicate the species complexes themselves have a much older origin, dating back perhaps to mid Miocene before the Baja peninsula broke off ~7 mya from the mainland at what is now Puerto Vallarta, Jalisco. Before then embayments and islands may have developed along the Pacific Coast of Mexico during 13�7 mya as �extension� of the �Proto-Gulf� occurred�which may be similar to the ancient seaway that developed in the Great Valley (Hall 2002; Brusca 2019; Spjut et al. 2020, Suppl. data file 1)�along with volcanic activity. The geology of Baja California is known for its volcanic fields as a result of the subduction of the Farallon Plate, 24�12 mya (Busby et al. 2020; Volcano Hotspot-agimarc 2019-03-17). Late Miocene alkali basalt-related lavas have been described along the Pacific Coast peninsula in northern Baja California for Mesa San Carlos and Mesa Santa Catarina, ~100 km south of El Rosario, to be capped by trachybasaltic (hawaiitic) flows dated between 9.3 and 7.5 mya (Pallares et al., 2007). They show multielement patterns of an ocean island basalt type (Luhr et al., 1995), including relatively recent activity at San Quint�n. The phylogeographic disjunction mentioned for many species complexes may also relate to the northward movement of the Baja California microplate that became attached to the Pacific Plate. Just glancing at Fig. 7 in Spjut et al. (2020), the Niebla specimen data in light blue, which are for specimens collected in the SVD, are generally positioned more basally. This is in contrast to SVD Vermilacinia specimens appearing more equally dispersed among the clades. Niebla generally occurs more inland than saxicolous Vermilacinia. This is because Niebla appears more adapted to receiving moisture from orographic fog as opposed to saxicolous Vermilacinia confined more to the coast, often with the ocean mist, and where humidity is higher with less change in diurnal temperature while also being able to absorb moisture from dew during the late evening to morning (Spjut 1996). With this in mind, Niebla populations are more affected by topographical and latitudinal change. Local occurrences of fog undoubtedly were impacted by frequent volcanic activity, causing a change in topography and wind patterns, relative to more gradual change in DNA by the northward movement of the Baja microplate, and even less so for those populations that remained at the same latitude. The following image shows the author standing on a slope inland where evidently orographic fog starts to precipitate as the moist air moves upslope, evident by the presence of Niebla at his feet but not evident beyond�towards the ocean.
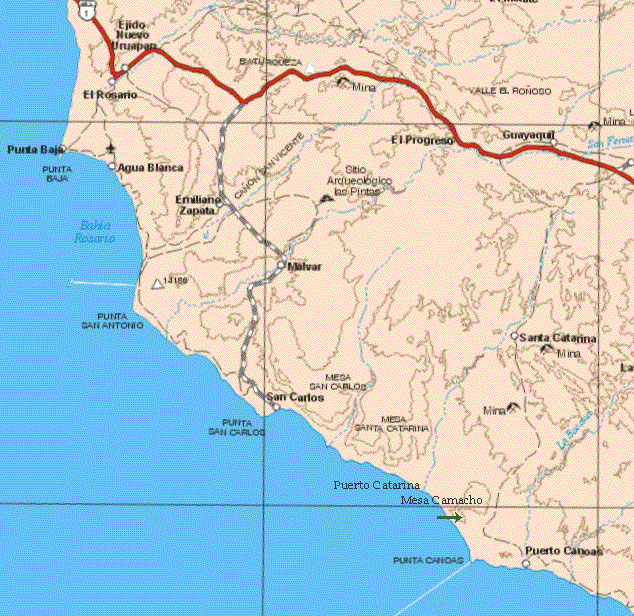
Cold upwelling water may have established along the Pacific Coast as early as late Oligocene in the San Gregorio Formation of Baja California Sur. This is evidenced by the association of organic-rich and finely laminated biosiliceous sediments with phosphorites (Grimm et al. 1991) even though coastal fog is generally associated with the California Current that evolved later (Kamikuri and Motoyama 2020). The occurrence of fog along the Pacific coast is related to upwelling of cold water coming into contact with the warmer atmosphere; the fog then moves inland due to differential atmospheric pressure and temperature between land and sea driven by offshore wind. This upwelling also shifts seasonally north from winter to summer (Ganeshram & Pedersen 1998; Goni et al. 2006). Since mid Miocene to early Pliocene (14�4.8 mya), a series of transitions occurred in the onshore-offshore location of West Coast upwelling (Barron & Bal-dauf 1990, Barron et al. 2002; Kamikuri & Motoyama 2020). Latitudinal position of the upwelling regime (Barron 1998), peaked both in its intensity and in surface area from 8 to 5 mya (Dickens & Owen 1999). During this time Baja California was nearly 300 km south of its present day position (Fletcher et al. 2007). As suggested for marine organisms along the Pacific Coast, species �radiated during peaks of upwelling primarily in the Late Miocene� and �secondarily in the Pleistocene� (Jacobs et al. 2004). Although phylogeny studies of Niebla do not support the taxonomy of Niebla species as circumscribed by Spjut (1996), their more restricted geographical range, nevertheless, suggests that all species can be supported at their type location. In this context, many species of Niebla are supported when phylogeny includes data on the type collection and excludes those specimens that are not within their phylogenetic clade. An example is six specimens identified Niebla lobulata for Spjut et al. (2020) that upon analysis by BPP indicated four species. Two of the six specimens are from the type locality (MD) shown in Fig. 7 (4861, 4866), and a third from the SVD (4812). They are monophyletic. Adding data from Jorna et al. (2021), seven specimens identified N. lobulata are monophyletic from the type locality, but Leavitt et al. also collected one from the MD (type locality), identified N. lobulata, that grouped with the N. fimbriata species complex in the CDT. Spjut et al. (2020) also identified N. lobulata collected in the CDT in association in the N. fimbriata complex, which in the ITS phylogenetic trees they appear in sister clades. Thus, N. lobulata reported to be polyphyletic by Spjut et al. (2020) is monophyletic sensu stricto, and not a micro-endemic, while it may also be interpreted to be phylogenetically related to cryptic species complexes, N. fimbriata and N. palmeri, when the type collection data are taken into consideration. Both studies focused on the overall taxonomy of Niebla based on circumscription of each of its species without addressing the results to type collection data; however, Spjut et al. (2020) pointed out that future phylogeny studies need to include type collection data. One difficulty is that many type locations are not easily accessible to obtain the fresh material that is required for DNA extraction, while there is no guarantee that the species will still be there. For example, Niebla suffnessii has vanished from Cerro El Elephante, a relatively isolated and undisturbed volcanic cone on the Vizca�no Peninsula north of Bah�a Asunci�n. The absence of N. suffnessii there in 2016 is obviously due to climate change; however, it was found just north of there on a lava mesa above Arroyo San Andr�s. This mesa is the type locality for the easily recognized N. usneoides (sekikaic acid) where observed to be common in 1986, and again in 1990, but N. usneoides could not be found in 2016, possibly because the main earth track around Cerro Elephante was once a more westerly loop nearer the ocean (further away from C. Elefante) as shown on the 1994 AAA map (Spjut 1996) compared with the its last issue published, 2003-04. Epitypification may be helpful for applying species names to phylogenetic studies, and if it is to be done, then it should be a Topotype that best fits the type chemistry and morphology, and possesses a genotype that is most phylogeographically related to the type collection based on a sampling of many specimens (see Lendemer 2020). Niebla reaches its greatest phenotypic diversity on peninsular Baja California between Punta San Carlos and Punta Rocosa. Six species of Niebla, and two species of Vermilacinia, were recognized endemic to this region. Other species of Niebla reach their southern distribution limits here such as Niebla homalea, generally not found south of Punta Baja except on Isla Cedros. Mesas between Punta Canoas and Puerto Catarina (including Mesa Camacho) are exceptionally rich in lichens, 28 species of Niebla were recorded from this region and include endemics such as Niebla tesselata, Vermilacinia vesiculosa, and three provisionally named (undescribed) species, N. angulata, N. sinuata and N. welwitschioides; see also Niebla and Vermilacinia communities of Baja California. Other Niebla rich communities occur further south on ridges above Punta Rocosa. The photo on the cover page shows a sandy ridge just south of Punta Negra where many type specimens were collected (Niebla flagelliforma, N. homaleoides, N. infundibula, N. juncosa, N. podetiaforma, N. sorocarpia, N. undulata, and Vermilacinia rigida). Because the DNA studies did resolve species of Niebla, or provide the data that is needed to corroborate or clarify the species, it seems best to retain the current taxonomy of the genus as published by Spjut (1996), in which the main clades based on chemistry are supported. A proposed modification is to recognize salazinic acid thalli of Niebla on the Vizca�no Peninsula as belonging to N. spatulata (species complex) instead of N. flabellata. |









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}